
1.1 Flower – A Fascinating
Organ of Angiosperms
1.2 Pre-fertilisation : Structures
and Events
1.3 Double Fertilisation
1.4 Post-fertilisation: Structures
and Events
1.5 Apomixis and
Polyembryony
Are we not lucky that plants reproduce sexually? The
myriads of flowers that we enjoy gazing at, the scents and
the perfumes that we swoon over, the rich colours that
attract us, are all there as an aid to sexual reproduction.
Flowers do not exist only for us to be used for our own
selfishness. All flowering plants show sexual reproduction.
A look at the diversity of structures of the inflorescences,
flowers and floral parts, shows an amazing range of
adaptations to ensure formation of the end products of
sexual reproduction, the fruits and seeds. In this chapter,
let us understand the morphology, structure and the
processes of sexual reproduction in flowering plants
(angiosperms).
1.1 FLOWER – A FASCINATING ORGAN OF
ANGIOSPERMS
Human beings have had an intimate relationship with
flowers since time immemorial. Flowers are objects of
aesthetic, ornamental, social, religious and cultural value
– they have always been used as symbols for conveying
important human feelings such as love, affection,
happiness, grief, mourning, etc. List at least five flowers
of ornamental value that are commonly cultivated at
homes and in gardens. Find out the names of five more flowers that are
used in social and cultural celebrations in your family. Have you heard
of floriculture – what does it refer to?
To a biologist, flowers are morphological and embryological marvels
and the sites of sexual reproduction. In earlier classes, you have read the
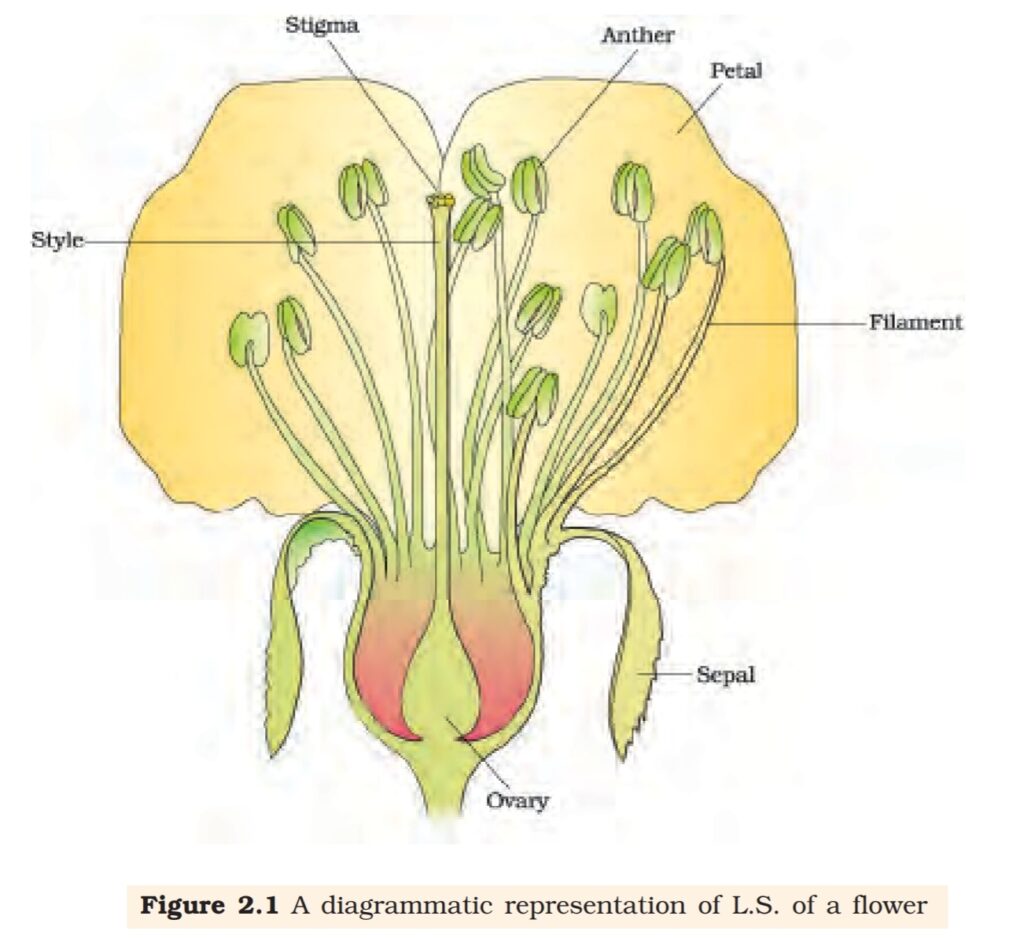
various parts of a flower. Figure 1.1 will help you recall the parts of a
typical flower. Can you name the two parts in a flower in which the two
most important units of sexual reproduction develop?
1.2 PRE-FERTILISATION: STRUCTURES AND EVENTS
Much before the actual flower is seen on a plant, the decision that the plant
is going to flower has taken place. Several hormonal and structural changes
are initiated which lead to the differentiation and further development of
the floral primordium. Inflorescences are formed which bear the floral buds
and then the flowers. In the flower the male and female reproductive
structures, the androecium and the gynoecium differentiate and develop.
You would recollect that the androecium consists of a whorl of stamens
representing the male reproductive organ and the gynoecium represents
the female reproductive organ.
1.2.1 Stamen, Microsporangium and Pollen Grain
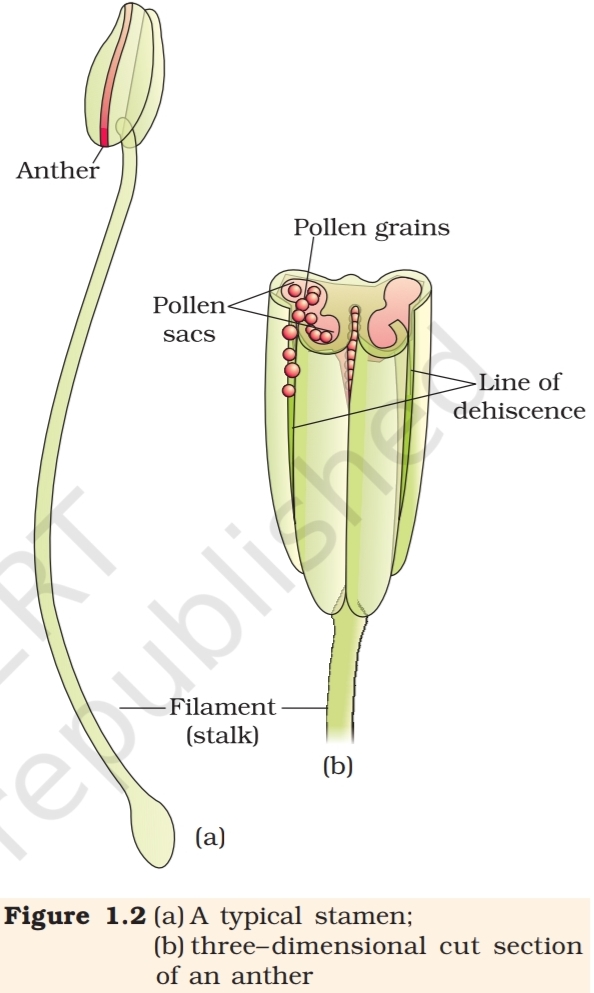
Figure 1.2a shows the two parts of a typical stamen – the long and slender
stalk called the filament, and the terminal generally bilobed structure
called the anther. The proximal end of the filament
is attached to the thalamus or the petal of the flower.
The number and length of stamens are variable in
flowers of different species. If you were to collect a
stamen each from ten flowers (each from different
species) and arrange them on a slide, you would
be able to appreciate the large variation in size seen
in nature. Careful observation of each stamen
under a dissecting microscope and making neat
diagrams would elucidate the range in shape and
attachment of anthers in different flowers.
A typical angiosperm anther is bilobed with
each lobe having two theca, i.e., they are dithecous
(Figure 1.2b). Often a longitudinal groove runs
lengthwise separating the theca. Let us
understand the various types of tissues and their
organisation in the transverse section of an anther
(Figure 1.3a). The bilobed nature of an anther is
very distinct in the transverse section of the anther.
The anther is a four-sided (tetragonal) structure
consisting of four microsporangia located at the
corners, two in each lobe.
The microsporangia develop further and
become pollen sacs. They extend longitudinally
all through the length of an anther and are packed
with pollen grains.
Structure of microsporangium: In a transverse
section, a typical microsporangium appears near
circular in outline. It is generally surrounded by four wall layers (Figure
1.3b)– the epidermis, endothecium, middle layers and the tapetum. The
outer three wall layers perform the function of protection and help in
dehiscence of anther to release the pollen. The innermost wall layer is
the tapetum. It nourishes the developing pollen grains. Cells of the
tapetum possess dense cytoplasm and generally have more than one
nucleus. Can you think of how tapetal cells could become bi-nucleate?
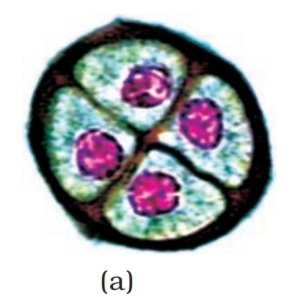
When the anther is young, a group of compactly arranged homogenous
cells called the sporogenous tissue occupies the centre of each
microsporangium.
Microsporogenesis : As the anther develops, the cells of the sporogenous
tissue undergo meiotic divisions to form microspore tetrads. What would
be the ploidy of the cells of the tetrad?

As each cell of the sporogenous tissue is capable of giving rise to a
microspore tetrad. Each one is a potential pollen or microspore mother
cell. The process of formation of microspores from a pollen mother cell (PMC)
through meiosis is called microsporogenesis. The microspores, as they
are formed, are arranged in a cluster of four cells–the microspore tetrad
(Figure 1.3a). As the anthers mature and dehydrate, the microspores
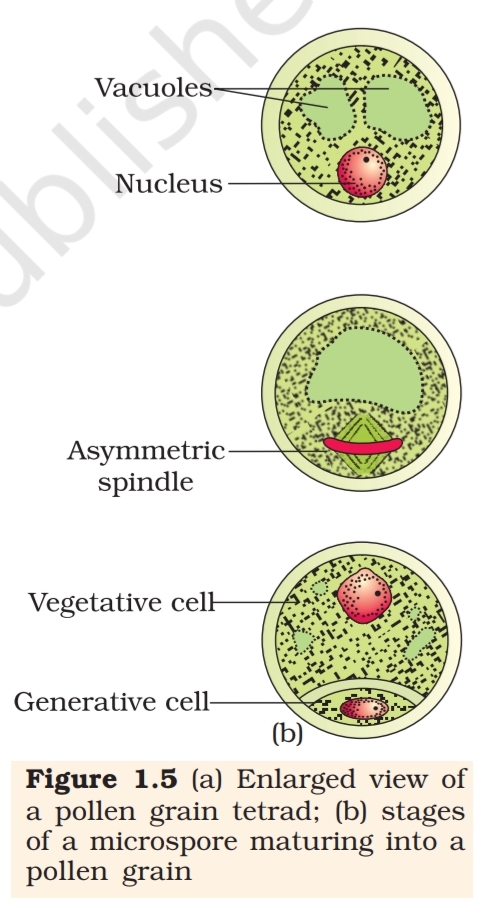
dissociate from each other and develop into pollen grains (Figure 1.3 b).
Inside each microsporangium several thousands of microspores or pollen
grains are formed that are released with the dehiscence of anther
(Figure 1.3c).
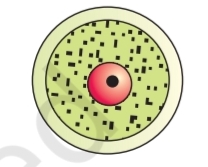
Pollen grain: The pollen grains represent the male gametophytes. If you
touch the opened anthers of Hibiscus or any other flower you would find
deposition of yellowish powdery pollen grains on your fingers. Sprinkle
these grains on a drop of water taken on a glass slide and observe under

a microscope. You will really be amazed at the variety of architecture –
sizes, shapes, colours, designs – seen on the pollen grains
from different species (Figure 1.4).
Pollen grains are generally spherical measuring about
25-50 micrometers in diameter. It has a prominent two-layered
wall. The hard outer layer called the exine is made up of
sporopollenin which is one of the most resistant organic material
known. It can withstand high temperatures and strong acids
and alkali. No enzyme that degrades sporopollenin is so far
known. Pollen grain exine has prominent apertures called germ
pores where sporopollenin is absent. Pollen grains are well-
preserved as fossils because of the presence of sporopollenin.
The exine exhibits a fascinating array of patterns and designs.
Why do you think the exine should be hard? What is the
function of germ pore? The inner wall of the pollen grain is
called the intine. It is a thin and continuous layer made up of
cellulose and pectin. The cytoplasm of pollen grain is
surrounded by a plasma membrane. When the pollen grain is
mature it contains two cells, the vegetative cell and generative
cell (Figure 1.5b). The vegetative cell is bigger, has abundant
food reserve and a large irregularly shaped nucleus. The
generative cell is small and floats in the cytoplasm of the
vegetative cell. It is spindle shaped with dense cytoplasm and
a nucleus. In over 60 per cent of angiosperms, pollen grains
are shed at this 2-celled stage. In the remaining species, the
generative cell divides mitotically to give rise to the two male
gametes before pollen grains are shed (3-celled stage).
Pollen grains of many species cause severe allergies and bronchial
afflictions in some people often leading to chronic respiratory
disorders– asthma, bronchitis, etc. It may be mentioned that Parthenium
or carrot grass that came into India as a contaminant with imported wheat,
has become ubiquitous in occurrence and causes pollen allergy.
Pollen grains are rich in nutrients. It has become a fashion in recent
years to use pollen tablets as food supplements. In western countries, a
large number of pollen products in the form of tablets and syrups are
available in the market. Pollen consumption has been claimed to increase
the performance of athletes and race horses (Figure 1.6).

When once they are shed, pollen grains have to land on the stigma
before they lose viability if they have to bring about fertilisation. How long
do you think the pollen grains retain viability? The period for which pollen
grains remain viable is highly variable and to some extent depends on the
prevailing temperature and humidity. In some cereals such as rice and
wheat, pollen grains lose viability within 30 minutes of their release, and
in some members of Rosaceae, Leguminoseae and Solanaceae, they
maintain viability for months. You may have heard of storing semen/
sperms of many animals including humans for artificial insemination. It
is possible to store pollen grains of a large number of species for years in
liquid nitrogen (-1960C). Such stored pollen can be used as pollen banks,
similar to seed banks, in crop breeding programmes.
1.2.2 The Pistil, Megasporangium (ovule) and Embryo sac
The gynoecium represents the female reproductive part of the flower. The
gynoecium may consist of a single pistil (monocarpellary) or may have
more than one pistil (multicarpellary). When there are more than one,
the pistils may be fused together (syncarpous) (Figure 1.7b) or may be
free (apocarpous) (Figure 1.7c). Each pistil has three parts (Figure 1.7a),
the stigma, style and ovary. The stigma serves as a landing platform
for pollen grains. The style is the elongated slender part beneath the
stigma. The basal bulged part of the pistil is the ovary. Inside the ovary
is the ovarian cavity (locule). The placenta is located inside the ovarian
cavity. Recall the definition and types of placentation that you studied in
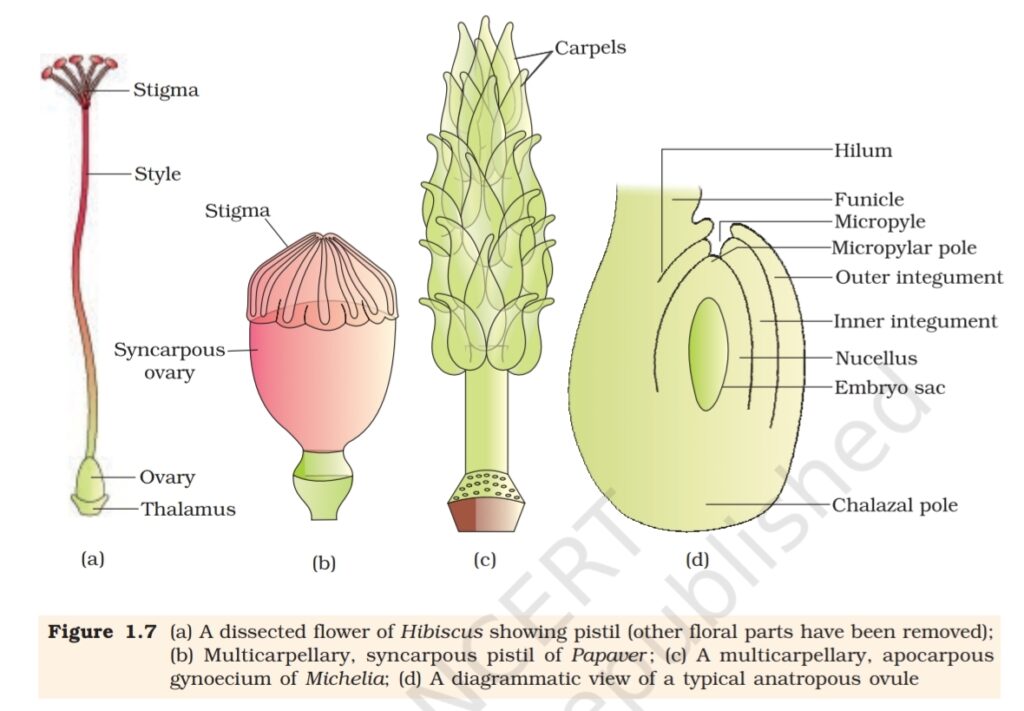
Class XI. Arising from the placenta are the megasporangia, commonly
called ovules. The number of ovules in an ovary may be one (wheat,
paddy, mango) to many (papaya, water melon, orchids).
The Megasporangium (Ovule) : Let us familiarise ourselves with the
structure of a typical angiosperm ovule (Figure 1.7d). The ovule is a small
structure attached to the placenta by means of a stalk called funicle.
The body of the ovule fuses with funicle in the region called hilum. Thus,
hilum represents the junction between ovule and funicle. Each ovule has
one or two protective envelopes called integuments. Integuments encircle
the nucellus except at the tip where a small opening called the micropyle
is organised. Opposite the micropylar end, is the chalaza, representing
the basal part of the ovule.
Enclosed within the integuments is a mass of cells called the nucellus.
Cells of the nucellus have abundant reserve food materials. Located in the
nucellus is the embryo sac or female gametophyte. An ovule generally has
a single embryo sac formed from a megaspore.
Megasporogenesis : The process of formation of megaspores from the
megaspore mother cell is called megasporogenesis. Ovules generally
differentiate a single megaspore mother cell (MMC) in the micropylar region

of the nucellus. It is a large cell containing dense cytoplasm and a
prominent nucleus. The MMC undergoes meiotic division. What is the
importance of the MMC undergoing meiosis? Meiosis results in the
production of four megaspores (Figure 1.8a).
Female gametophyte : In a majority of flowering plants, one of the
megaspores is functional while the other three degenerate. Only the
functional megaspore develops into the female gametophyte (embryo
sac). This method of embryo sac formation from a single megaspore is termed
monosporic development. What will be the ploidy of the cells of the nucellus,
MMC, the functional megaspore and female gametophyte?
Let us study about the formation of the embryo sac in detail.
(Figure 1.8b). The nucleus of the functional megaspore divides mitotically
to form two nuclei which move to the opposite poles, forming the 2-
nucleate embryo sac. Two more sequential mitotic nuclear divisions
result in the formation of the 4-nucleate and later the 8-nucleate stages
of the embryo sac. It is of interest to note that these mitotic divisions are
strictly free nuclear, that is, nuclear divisions are not followed immediately
by cell wall formation. After the 8-nucleate stage, cell walls are laid down
leading to the organisation of the typical female gametophyte
or embryo sac. Observe the distribution of cells inside the embryo sac
(Figure 1.8b, c). Six of the eight nuclei are surrounded by cell walls and
organised into cells; the remaining two nuclei, called polar nuclei are
situated below the egg apparatus in the large central cell.
There is a characteristic distribution of the cells within the embryo
sac. Three cells are grouped together at the micropylar end and constitute
the egg apparatus. The egg apparatus, in turn, consists of two synergids
and one egg cell. The synergids have special cellular thickenings at the
micropylar tip called filiform apparatus, which play an important role in
guiding the pollen tubes into the synergid. Three cells are at the chalazal
end and are called the antipodals. The large central cell, as mentioned
earlier, has two polar nuclei. Thus, a typical angiosperm embryo sac, at
maturity, though 8-nucleate is 7-celled.
1.2.3 Pollination
In the preceding sections you have learnt that the male and female gametes
in flowering plants are produced in the pollen grain and embryo sac,
respectively. As both types of gametes are non-motile, they have to be
brought together for fertilisation to occur. How is this achieved?
Pollination is the mechanism to achieve this objective. Transfer
of pollen grains (shed from the anther) to the stigma of a pistil is
termed pollination. Flowering plants have evolved an amazing array
of adaptations to achieve pollination. They make use of external
agents to achieve pollination. Can you list the possible external
agents?
Kinds of Pollination : Depending on the source of pollen, pollination
can be divided into three types.
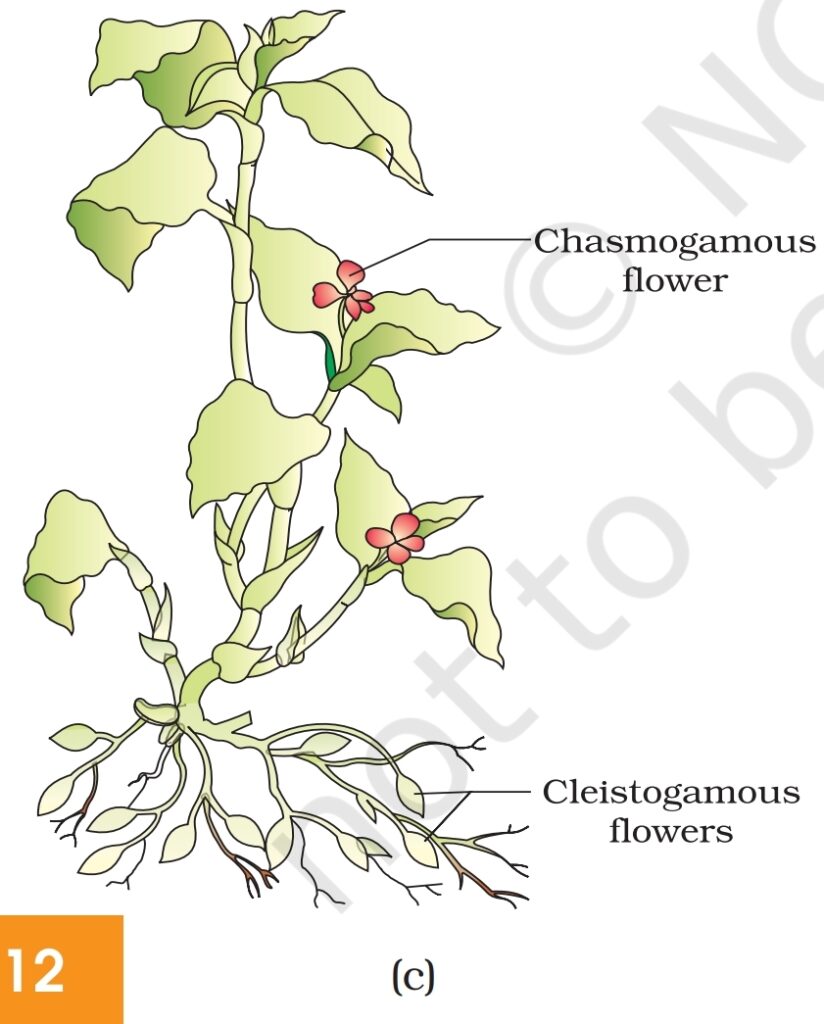
(i) Autogamy : In this type, pollination is achieved within the same
flower. Transfer of pollen grains from the anther to the stigma of the
same flower (Figure 1.9a). In a normal flower which opens and
exposes the anthers and the stigma, complete autogamy is rather
rare. Autogamy in such flowers requires synchrony in pollen release
and stigma receptivity and also, the anthers and the stigma should
lie close to each other so that self-pollination
can occur. Some plants such as Viola
(common pansy), Oxalis, and Commelina
produce two types of flowers –
chasmogamous flowers which are similar to
flowers of other species with exposed anthers
and stigma, and cleistogamous flowers which
do not open at all (Figure 1.9c). In such flowers,
the anthers and stigma lie close to each other.
When anthers dehisce in the flower buds,
pollen grains come in contact with the stigma
to effect pollination. Thus, cleistogamous
flowers are invariably autogamous as there is
no chance of cross-pollen landing on the
stigma. Cleistogamous flowers produce
assured seed-set even in the absence of
pollinators. Do you think that cleistogamy is
advantageous or disadvantageous to the
plant? Why?



(ii) Geitonogamy – Transfer of pollen grains from
the anther to the stigma of another flower of
the same plant. Although geitonogamy is
functionally cross-pollination involving a
pollinating agent, genetically it is similar to
autogamy since the pollen grains come from
the same plant.
(iii) Xenogamy – Transfer of pollen grains from
anther to the stigma of a different plant (Figure
1.9b). This is the only type of pollination which
during pollination brings genetically different
types of pollen grains to the stigma.
Agents of Pollination : Plants use two abiotic (wind
and water) and one biotic (animals) agents to achieve
pollination. Majority of plants use biotic agents for
pollination. Only a small proportion of plants use
abiotic agents. Pollen grains coming in contact with
the stigma is a chance factor in both wind and water
pollination. To compensate for this uncertainties and
associated loss of pollen grains, the flowers produce
enormous amount of pollen when compared to the
number of ovules available for pollination.
Pollination by wind is more common
amongst abiotic pollinations. Wind pollination
also requires that the pollen grains are light
and non-sticky so that they can be
transported in wind currents. They often
possess well-exposed stamens (so that the
pollens are easily dispersed into wind currents,
Figure 1.10) and large often-feathery stigma
to easily trap air-borne pollen grains. Wind-
pollinated flowers often have a single ovule in
each ovary and numerous flowers packed into
an inflorescence; a familiar example is the corn
cob – the tassels you see are nothing but the
stigma and style which wave in the wind to
trap pollen grains. Wind-pollination is quite
common in grasses.

Pollination by water is quite rare in
flowering plants and is limited to about 30
genera, mostly monocotyledons. As against
this, you would recall that water is a regular
mode of transport for the male gametes among
the lower plant groups such as algae,
bryophytes and pteridophytes. It is believed,
particularly for some bryophytes and
pteridophytes, that their distribution is limited
because of the need for water for the transport
of male gametes and fertilisation. Some
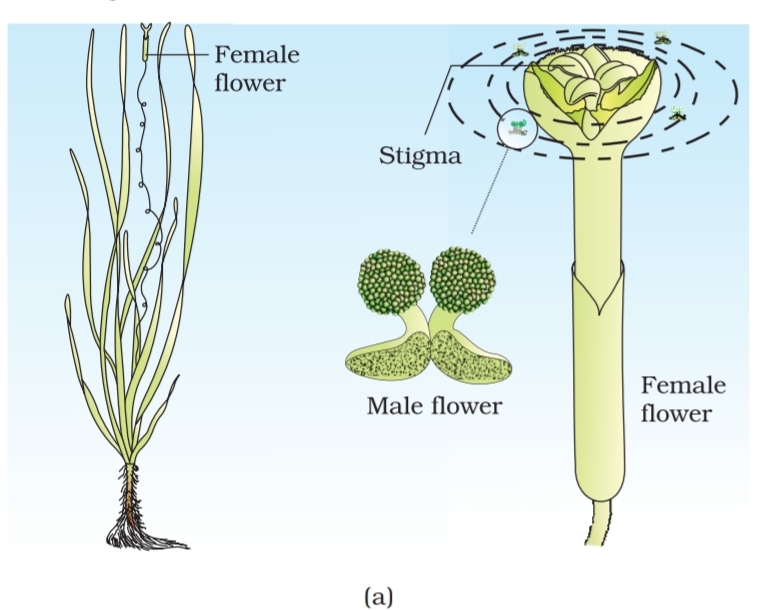
examples of water pollinated plants are Vallisneria and Hydrilla which
grow in fresh water and several marine sea-grasses such as Zostera. Not
all aquatic plants use water for pollination. In a majority of aquatic plants
such as water hyacinth and water lily, the flowers emerge above the level
of water and are pollinated by insects or wind as in most of the land
plants. In Vallisneria, the female flower reach the surface of water by the
long stalk and the male flowers or pollen grains are released on to the
surface of water. They are carried passively by water currents (Figure
1.11a); some of them eventually reach the female flowers and the stigma.
In another group of water pollinated plants such as seagrasses, female
flowers remain submerged in water and the pollen grains are released
inside the water. Pollen grains in many such species are long, ribbon like
and they are carried passively inside the water; some of them reach the
stigma and achieve pollination. In most of the water-pollinated species,
pollen grains are protected from wetting by a mucilaginous covering.
Both wind and water pollinated flowers are not very colourful and do
not produce nectar. What would be the reason for this?
Majority of flowering plants use
a range of animals as pollinating
agents. Bees, butterflies, flies,
beetles, wasps, ants, moths, birds
(sunbirds and humming birds) and
bats are the common pollinating
agents. (Figure 1.11b). Among the
animals, insects, particularly bees
are the dominant biotic pollinating
agents. Even larger animals such
as some primates (lemurs), arboreal
(tree-dwelling) rodents, or even
reptiles (gecko lizard and garden
lizard) have also been reported as
pollinators in some species.
Often flowers of animal-
pollinated plants are specifically
adapted for a particular species of
animal.


Majority of insect-pollinated
flowers are large, colourful, fragrant
and rich in nectar. When the flowers
are small, a number of flowers are
clustered into an inflorescence to
make them conspicuous. Animals
are attracted to flowers by colour
and/or fragrance. The flowers
pollinated by flies and beetles
secrete foul odours to attract these
animals. To sustain animal visits,
the flowers have to provide rewards
to the animals. Nectar and pollen
grains are the usual floral rewards.
For harvesting the reward(s) from
the flower the animal visitor comes
in contact with the anthers and the
stigma. The body of the animal gets
a coating of pollen grains, which are
generally sticky in animal pollinated flowers. When the animal carrying
pollen on its body comes in contact with the stigma, it brings about
pollination.
In some species floral rewards are in providing safe places to lay eggs;
an example is that of the tallest flower of Amorphophallus (the flower
itself is about 6 feet in height). A similar relationship exists between a
species of moth and the plant Yucca where both species – moth and the
plant – cannot complete their life cycles without each other. The moth
deposits its eggs in the locule of the ovary and the flower, in turn, gets
pollinated by the moth. The larvae of the moth come out of the eggs as
the seeds start developing.
Why don’t you observe some flowers of the following plants (or any
others available to you): Cucumber, Mango, Peepal, Coriander, Papaya,
Onion, Lobia, Cotton, Tobacco, Rose, Lemon, Eucalyptus, Banana? Try to
find out which animals visit them and whether they could be
pollinators.You’ll have to patiently observe the flowers over a few days
and at different times of the day. You could also try to see whether there
is any correlation in the characteristics of a flower to the animal that
visits it. Carefully observe if any of the visitors come in contact with the
anthers and the stigma as only such visitors can bring about pollination.
Many insects may consume pollen or the nectar without bringing about
pollination. Such floral visitors are referred to as pollen/nectar robbers.
You may or may not be able to identify the pollinators, but you will surely
enjoy your efforts!
Outbreeding Devices : Majority of flowering plants produce hermaphrodite
flowers and pollen grains are likely to come in contact with the stigma of
the same flower. Continued self-pollination result in inbreeding depression.
Flowering plants have developed many devices to discourage self-
pollination and to encourage cross-pollination. In some species, pollen
release and stigma receptivity are not synchronised. Either the pollen is
released before the stigma becomes receptive or stigma becomes receptive
much before the release of pollen. In some other species, the anther and
stigma are placed at different positions so that the pollen cannot come in
contact with the stigma of the same flower. Both these devices prevent
autogamy. The third device to prevent inbreeding is self-incompatibility.
This is a genetic mechanism and prevents self-pollen (from the same flower
or other flowers of the same plant) from fertilising the ovules by inhibiting
pollen germination or pollen tube growth in the pistil. Another device to
prevent self-pollination is the production of unisexual flowers. If both male
and female flowers are present on the same plant such as castor and maize
(monoecious), it prevents autogamy but not geitonogamy. In several species
such as papaya, male and female flowers are present on different plants,
that is each plant is either male or female (dioecy). This condition prevents
both autogamy and geitonogamy.
Pollen-pistil Interaction : Pollination does not guarantee the transfer
of the right type of pollen (compatible pollen of the same species as the
stigma). Often, pollen of the wrong type, either from other species or from
the same plant (if it is self-incompatible), also land on the stigma. The
pistil has the ability to recognise the pollen, whether it is of the right type
(compatible) or of the wrong type (incompatible). If it is of the right type,
the pistil accepts the pollen and promotes post-pollination events that

leads to fertilisation. If the pollen is of the wrong type, the pistil rejects the
pollen by preventing pollen germination on the stigma or the pollen tube
growth in the style. The ability of the pistil to recognise the pollen followed
by its acceptance or rejection is the result of a continuous dialogue
between pollen grain and the pistil. This dialogue is mediated by chemical
components of the pollen interacting with those of the pistil. It is only in
recent years that botanists have been able to identify some of the pollen
and pistil components and the interactions leading to the recognition,
followed by acceptance or rejection.
As mentioned earlier, following compatible pollination, the pollen grain
germinates on the stigma to produce a pollen tube through one of the
germ pores (Figure 1.12a). The contents of the pollen grain move into the
pollen tube. Pollen tube grows through the tissues of the stigma and
style and reaches the ovary (Figure 1.12b, c). You would recall that in
some plants, pollen grains are shed at two-celled condition (a vegetative
cell and a generative cell). In such plants, the generative cell divides and
forms the two male gametes during the growth of pollen tube in the stigma.
In plants which shed pollen in the three-celled condition, pollen tubes
carry the two male gametes from the beginning. Pollen tube, after reaching
the ovary, enters the ovule through the micropyle and then enters one of
the synergids through the filiform apparatus (Figure 1.12d, e). Many recent
studies have shown that filiform apparatus present at the micropylar part
of the synergids guides the entry of pollen tube. All these events–from
pollen deposition on the stigma until pollen tubes enter the ovule–are
together referred to as pollen-pistil interaction. As pointed out earlier,
pollen-pistil interaction is a dynamic process involving pollen recognition
followed by promotion or inhibition of the pollen. The knowledge gained
in this area would help the plant breeder in manipulating pollen-pistil
interaction, even in incompatible pollinations, to get desired hybrids.
You can easily study pollen germination by dusting some pollen from
flowers such as pea, chickpea, Crotalaria, balsam and Vinca on a glass slide
containing a drop of sugar solution (about 10 per cent). After about 15–30
minutes, observe the slide under the low power lens of the microscope. You
are likely to see pollen tubes coming out of the pollen grains.
A breeder is interested in crossing different species and often genera
to combine desirable characters to produce commercially ‘superior’
varieties. Artificial hybridisation is one of the major approaches of crop
improvement programme. In such crossing experiments it is important
to make sure that only the desired pollen grains are used for pollination
and the stigma is protected from contamination (from unwanted pollen).
This is achieved by emasculation and bagging techniques.
If the female parent bears bisexual flowers, removal of anthers from
the flower bud before the anther dehisces using a pair of forceps is
necessary. This step is referred to as emasculation. Emasculated flowers
have to be covered with a bag of suitable size, generally made up of butter
paper, to prevent contamination of its stigma with unwanted pollen. This
process is called bagging. When the stigma of bagged flower attains
receptivity, mature pollen grains collected from anthers of the male parent
are dusted on the stigma, and the flowers are rebagged, and the fruits
allowed to develop.
If the female parent produces unisexual flowers, there is no need for
emasculation. The female flower buds are bagged before the flowers open.
When the stigma becomes receptive, pollination is carried out using the
desired pollen and the flower rebagged.
Ration
1.3 DOUBLE FERTILISATION
After entering one of the synergids, the pollen tube releases the two male
gametes into the cytoplasm of the synergid. One of the male gametes moves
towards the egg cell and fuses with its nucleus thus completing the
syngamy. This results in the formation of a diploid cell, the zygote. The
other male gamete moves towards the two polar nuclei located in the central
cell and fuses with them to produce a triploid primary endosperm nucleus
(PEN) (Figure 1.13a). As this involves the fusion of three haploid nuclei it
is termed triple fusion. Since two types of fusions, syngamy and triple
fusion take place in an embryo sac the phenomenon is termed double
fertilisation, an event unique to flowering plants. The central cell after
triple fusion becomes the primary endosperm cell (PEC) and develops
into the endosperm while the zygote develops into an embryo.

1.4 POST-FERTILISATION : STRUCTURES AND EVENTS
Following double fertilisation, events of endosperm and embryo
development, maturation of ovule(s) into seed(s) and ovary into fruit, are
collectively termed post-fertilisation events.
1.4.1 Endosperm
Endosperm development precedes embryo development. Why? The
primary endosperm cell divides repeatedly and forms a triploid
endosperm tissue. The cells of this tissue are filled with
reserve food materials and are used for the nutrition of
the developing embryo. In the most common type of
endosperm development, the PEN undergoes successive
nuclear divisions to give rise to free nuclei. This stage of
endosperm development is called free-nuclear endosperm.
Subsequently cell wall formation occurs and the
endosperm becomes cellular. The number of free nuclei
formed before cellularisation varies greatly. The coconut
water from tender coconut that you are familiar with, is
nothing but free-nuclear endosperm (made up of
thousands of nuclei) and the surrounding white kernel is
the cellular endosperm.
Endosperm may either be completely consumed by the
developing embryo (e.g., pea, groundnut, beans) before seed
maturation or it may persist in the mature seed (e.g. castor
and coconut) and be used up during seed germination. Split
open some seeds of castor, peas, beans, groundnut, fruit of
coconut and look for the endosperm in each case. Find out
whether the endosperm is persistent in cereals – wheat, rice
and maize.
1.4.2 Embryo
Embryo develops at the micropylar end of the embryo sac where
the zygote is situated. Most zygotes divide only after certain
amount of endosperm is formed. This is an adaptation to
provide assured nutrition to the developing embryo. Though
the seeds differ greatly, the early stages of embryo development
(embryogeny) are similar in both monocotyledons and
dicotyledons. Figure 1.13 depicts the stages of embryogeny in
a dicotyledonous embryo. The zygote gives rise to the
proembryo and subsequently to the globular, heart-shaped
and mature embryo.
A typical dicotyledonous embryo (Figure 1.14a), consists
of an embryonal axis and two cotyledons. The portion of
embryonal axis above the level of cotyledons is the epicotyl,
which terminates with the plumule or stem tip. The cylindrical
portion below the level of cotyledons is hypocotyl that
terminates at its lower end in the radicle or root tip. The root
tip is covered with a root cap.
Embryos of monocotyledons (Figure 1.14 b) possess only
one cotyledon. In the grass family the cotyledon is called
scutellum that is situated towards one side (lateral) of the
embryonal axis. At its lower end, the embryonal axis has the

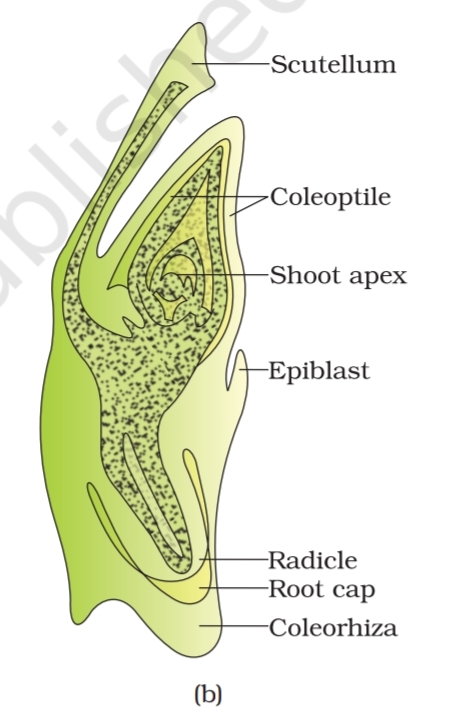

radical and root cap enclosed in an undifferentiated sheath called
coleorrhiza. The portion of the embryonal axis above the level of
attachment of scutellum is the epicotyl. Epicotyl has a shoot apex and a
few leaf primordia enclosed in a hollow foliar structure, the coleoptile.
Soak a few seeds in water (say of wheat, maize, peas, chickpeas,
ground nut) overnight. Then split the seeds and observe the various
parts of the embryo and the seed.
1.4.3 Seed
In angiosperms, the seed is the final product of sexual reproduction. It is
often described as a fertilised ovule. Seeds are formed inside fruits. A
seed typically consists of seed coat(s), cotyledon(s) and an embryo axis.
The cotyledons (Figure 1.15a) of the embryo are simple structures,
generally thick and swollen due to storage of food reserves (as in legumes).
Mature seeds may be non-albuminous or ex-albuminous. Non-
albuminous seeds have no residual endosperm as it is completely
consumed during embryo development (e.g., pea, groundnut).
Albuminous seeds retain a part of endosperm as it is not completely used
up during embryo development (e.g., wheat, maize, barley, castor).
Occasionally, in some seeds such as black pepper and beet, remnants of
nucellus are also persistent. This residual, persistent nucellus is the
perisperm.
Integuments of ovules harden as tough protective seed coats
(Figure 1.15a). The micropyle remains as a small pore in the seed coat.
This facilitates entry of oxygen and water into the seed during germination.
As the seed matures, its water content is reduced and seeds become
relatively dry (10-15 per cent moisture by mass). The general metabolic
activity of the embryo slows down. The embryo may enter a state of
inactivity called dormancy, or if favourable conditions are available
(adequate moisture, oxygen and suitable temperature), they germinate.
As ovules mature into seeds, the ovary develops into a fruit, i.e., the
transformation of ovules into seeds and ovary into fruit proceeds
simultaneously. The wall of the ovary develops into the wall of fruit called
pericarp. The fruits may be fleshy as in guava, orange, mango, etc., or
may be dry, as in groundnut, and mustard, etc. Many fruits have evolved
mechanisms for dispersal of seeds. Recall the classification of fruits and
their dispersal mechanisms that you have studied in an earlier class. Is
there any relationship between number of ovules in an ovary and the
number of seeds present in a fruit?
In most plants, by the time the fruit develops from the ovary, other
floral parts degenerate and fall off. However, in a few species such as apple,
strawberry, cashew, etc., the thalamus also contributes to fruit formation.
Such fruits are called false fruits (Figure 1.15b). Most fruits however
develop only from the ovary and are called true fruits. Although in most
of the species, fruits are the results of fertilisation, there are a few species
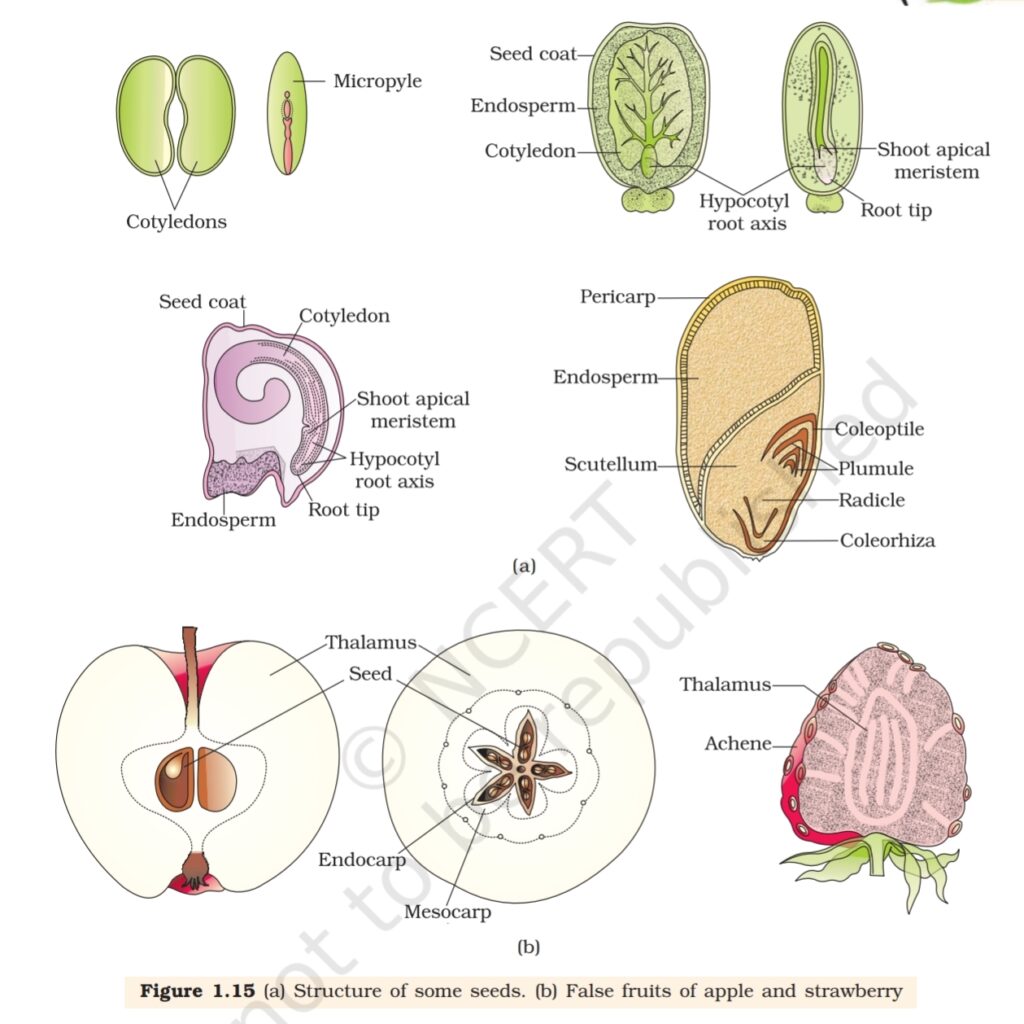
in which fruits develop without fertilisation. Such fruits are called
parthenocarpic fruits. Banana is one such example. Parthenocarpy can
be induced through the application of growth hormones and such fruits
are seedless.
Seeds offer several advantages to angiosperms. Firstly, since
reproductive processes such as pollination and fertilisation are
independent of water, seed formation is more dependable. Also seeds have
better adaptive strategies for dispersal to new habitats and help the species
to colonise in other areas. As they have sufficient food reserves, young
seedlings are nourished until they are capable of photosynthesis on their
own. The hard seed coat provides protection to the young embryo. Being
products of sexual reproduction, they generate new genetic combinations
leading to variations.
Seed is the basis of our agriculture. Dehydration and dormancy of
mature seeds are crucial for storage of seeds which can be used as food
throughout the year and also to raise crop in the next season. Can you
imagine agriculture in the absence of seeds, or in the presence of seeds
which germinate straight away soon after formation and cannot be stored?
How long do the seeds remain alive after they are dispersed? This
period again varies greatly. In a few species the seeds lose viability within
a few months. Seeds of a large number of species live for several years.
Some seeds can remain alive for hundreds of years. There are several
records of very old yet viable seeds. The oldest is that of a lupine, Lupinus
arcticus excavated from Arctic Tundra. The seed germinated and flowered
after an estimated record of 10,000 years of dormancy. A recent record of
2000 years old viable seed is of the date palm, Phoenix dactylifera
discovered during the archeological excavation at King Herod’s palace
near the Dead Sea.
After completing a brief account of sexual reproduction of flowering
plants it would be worth attempting to comprehend the enormous
reproductive capacity of some flowering plants by asking the following
questions: How many eggs are present in an embryo sac? How many
embryo sacs are present in an ovule? How many ovules are present in
an ovary? How many ovaries are present in a typical flower? How many
flowers are present on a tree? And so on…
Can you think of some plants in which fruits contain very large
number of seeds. Orchid fruits are one such category and each fruit
contain thousands of tiny seeds. Similar is the case in fruits of some
parasitic species such as Orobanche and Striga. Have you seen a tiny
seed of Ficus? How large is the tree of Ficus developed from that tiny
seed. How many billions of seeds does each Ficus tree produce? Can
you imagine any other example in which such a tiny structure can
produce such a large biomass over the years?
1.5 APOMIXIS AND POLYEMBRYONY
Although seeds, in general are the products of fertilisation, a few flowering
plants such as some species of Asteraceae and grasses, have evolved a
special mechanism, to produce seeds without fertilisation, called apomixis.
What is fruit production without fertilisation called? Thus, apomixis is a
form of asexual reproduction that mimics sexual reproduction. There are
several ways of development of apomictic seeds. In some species, the
diploid egg cell is formed without reduction division and develops into
the embryo without fertilisation. More often, as in many Citrus and Mango
varieties some of the nucellar cells surrounding the embryo sac start
dividing, protrude into the embryo sac and develop into the embryos. In
such species each ovule contains many embryos. Occurrence of more
than one embryo in a seed is referred to as polyembryony. Take out
some seeds of orange and squeeze them. Observe the many embryos of
different sizes and shapes from each seed. Count the number of embryos
in each seed. What would be the genetic nature of apomictic embryos?
Can they be called clones?
Hybrid varieties of several of our food and vegetable crops are being
extensively cultivated. Cultivation of hybrids has tremendously increased
productivity. One of the problems of hybrids is that hybrid seeds have
to be produced every year. If the seeds collected from hybrids are sown,
the plants in the progeny will segregate and do not maintain hybrid
characters. Production of hybrid seeds is costly and hence the cost of
hybrid seeds become too expensive for the farmers. If these hybrids are
made into apomicts, there is no segregation of characters in the hybrid
progeny. Then the farmers can keep on using the hybrid seeds to raise
new crop year after year and he does not have to buy hybrid seeds every
year. Because of the importance of apomixis in hybrid seed industry,
active research is going on in many laboratories around the world to
understand the genetics of apomixis and to transfer apomictic genes
into hybrid varieties.
SUMMARY
Flowers are the seat of sexual reproduction in angiosperms. In the flower,
androecium consisting of stamens represents the male reproductive
organs and gynoecium consisting of pistils represents the female
reproductive organs.
A typical anther is bilobed, dithecous and tetrasporangiate. Pollen
grains develop inside the microsporangia. Four wall layers, the
epidermis, endothecium, middle layers and the tapetum surround the
microsporangium. Cells of the sporogenous tissue lying in the centre of
the microsporangium, undergo meiosis (microsporogenesis) to form
tetrads of microspores. Individual microspores mature into pollen grains.
Pollen grains represents the male gametophytic generation. The
pollen grains have a two-layered wall, the outer exine and inner intine.
The exine is made up of sporopollenin and has germ pores. Pollen grains
may have two cells (a vegetative cell and generative cell) or three cells (a
vegetative cell and two male gametes) at the time of shedding.
The pistil has three parts – the stigma, style and the ovary. Ovules
are present in the ovary. The ovules have a stalk called funicle, protective
integument(s), and an opening called micropyle. The central tissue is
the nucellus in which the archesporium differentiates. A cell of the
archesporium, the megaspore mother cell divides meiotically and one of
the megaspores forms the embryo sac (the female gametophyte). The
mature embryo sac is 7-celled and 8-nucleate. At the micropylar end is
the egg apparatus consisting of two synergids and an egg cell. At the
chalazal end are three antipodals. At the centre is a large central cell
with two polar nuclei.
Pollination is the mechanism to transfer pollen grains from the
anther to the stigma. Pollinating agents are either abiotic (wind and
water) or biotic (animals).
Pollen-pistil interaction involves all events from the landing of pollen
grains on the stigma until the pollen tube enters the embryo sac (when
the pollen is compatible) or pollen inhibition (when the pollen is
incompatible). Following compatible pollination, pollen grain germinates
on the stigma and the resulting pollen tube grow through the style,
enter the ovules and finally discharges two male gametes in one of the
synergids. Angiosperms exhibit double fertilisation because two fusion
events occur in each embryo sac, namely syngamy and triple fusion.
The products of these fusions are the diploid zygote and the triploid
primary endosperm nucleus (in the primary endosperm cell). Zygote
develops into the embryo and the primary endosperm cell forms the
endosperm tissue. Formation of endosperm always precedes
development of the embryo.
The developing embryo passes through different stages such as
the proembryo, globular and heart-shaped stages before maturation.
Mature dicotyledonous embryo has two cotyledons and an embryonal
axis with epicotyl and hypocotyl. Embryos of monocotyledons have a
single cotyledon. After fertilisation, ovary develops into fruit and ovules
develop into seeds.
A phenomenon called apomixis is found in some angiosperms,
particularly in grasses. It results in the formation of seeds without
fertilisation. Apomicts have several advantages in horticulture and
agriculture.
Some angiosperms produce more than one embryo in their seed.
This phenomenon is called polyembryony.
EXERCISES
- Name the parts of an angiosperm flower in which development of male
and female gametophyte take place. - Differentiate between microsporogenesis and megasporogenesis. Which
type of cell division occurs during these events? Name the structures
formed at the end of these two events. - Arrange the following terms in the correct developmental sequence:
Pollen grain, sporogenous tissue, microspore tetrad, pollen mother cell,
male gametes. - With a neat, labelled diagram, describe the parts of a typical angiosperm
ovule. - What is meant by monosporic development of female gametophyte?
- With a neat diagram explain the 7-celled, 8-nucleate nature of the female
gametophyte. - What are chasmogamous flowers? Can cross-pollination occur in
cleistogamous flowers? Give reasons for your answer. - Mention two strategies evolved to prevent self-pollination in flowers.
- What is self-incompatibility? Why does self-pollination not lead to seed
formation in self-incompatible species? - What is bagging technique? How is it useful in a plant breeding
programme? - What is triple fusion? Where and how does it take place? Name the
nuclei involved in triple fusion. - Why do you think the zygote is dormant for sometime in a fertilised
ovule? - Differentiate between:
(a) hypocotyl and epicotyl;
(b) coleoptile and coleorrhiza;
(c) integument and testa;
(d) perisperm and pericarp. - Why is apple called a false fruit? Which part(s) of the flower forms the
fruit? - What is meant by emasculation? When and why does a plant breeder
employ this technique? - If one can induce parthenocarpy through the application of growth
substances, which fruits would you select to induce parthenocarpy
and why? - Explain the role of tapetum in the formation of pollen-grain wall.
- What is apomixis and what is its importance?
507 responses to “CHAPTER 2 SEXUAL REPRODUCTION INFLOWERING PLANTS”
Please tell me more about your excellent articles
Thank you for writing this post!
What’s Happening i’m new to this, I stumbled upon this I have found It positively useful and it has helped me out loads. I hope to contribute & assist other users like its aided me. Good job.
Thanks for posting. I really enjoyed reading it, especially because it addressed my problem. It helped me a lot and I hope it will help others too.
Thanks for your help and for writing this post. It’s been great.
Thank you for writing this article. I appreciate the subject too.
Please provide me with more details on the topic
If you appreciate the insights presented in this blog post, consider subscribing for future updates. Staying informed is crucial in our ever-evolving landscapevisit sexy
The quality of this content is commendable; it not only informs but also stimulates critical thinking among readers. Such diligence in research is greatly appreciated sexy
I appreciate the effort put into creating this content. It reflects a high standard of professionalism and contributes meaningfully to ongoing discussions in the field sexy
The quality of this content is commendable; it not only informs but also stimulates critical thinking among readers. Such diligence in research is greatly appreciated sexy
Wow, what an engaging read I found the insights in this article to be incredibly thought-provoking. Thanks for sharing the link sexy
Such a very interesting article It sparked so many ideas in my mind. Thank you for sharing this gem with us sexy
Such a very interesting article It sparked so many ideas in my mind. Thank you for sharing this gem with us sexy
Wow, what an engaging read I found the insights in this article to be incredibly thought-provoking. Thanks for sharing the link sexy
If you found this blog post insightful, I encourage you to explore further resources on the topic. Your engagement with such content contributes to a deeper understanding of the subject mattervisit sexy
If this blog post aligns with your interests, I invite you to reflect on its implications and share your thoughts in the comments section. Engaging in dialogue enriches our comprehension of complex issuesvisit sexy
This article is so fascinating I really appreciate how it breaks down complex ideas into easily digestible parts. Can’t wait to read more from this site sexy
What a very interesting article I loved how it delved into the nuances of the subject. Thanks for sharing this link sexy
This article is so fascinating I really appreciate how it breaks down complex ideas into easily digestible parts. Can’t wait to read more from this site sexy
I really enjoyed this article It’s amazing how much depth and detail you covered. Looking forward to exploring more on your site sexy
If you found value in this blog post, please consider expressing your support through comments or shares. Contributions from readers like yourself are vital for nurturing an informed communityvisit sexy
This article is so fascinating I really appreciate how it breaks down complex ideas into easily digestible parts. Can’t wait to read more from this site sexy
I really enjoyed this article It’s amazing how much depth and detail you covered. Looking forward to exploring more on your site sexy
Hi there! Do you know if they make any plugins to help with SEO?
I’m trying to get my site to rank for some targeted keywords but I’m not
seeing very good results. If you know of any please share.
Many thanks! You can read similar art here:
Bij nl
I commend the author for presenting such high-quality content. The clarity and depth of analysis truly enhance the reader’s experience sexy
If you found this blog post insightful, I encourage you to explore further resources on the topic. Your engagement with such content contributes to a deeper understanding of the subject mattervisit sexy
I really enjoyed this article It’s amazing how much depth and detail you covered. Looking forward to exploring more on your site sexy
I commend the author for presenting such high-quality content. The clarity and depth of analysis truly enhance the reader’s experience sexy
Wow, what an engaging read I found the insights in this article to be incredibly thought-provoking. Thanks for sharing the link sexy
I really enjoyed this article It’s amazing how much depth and detail you covered. Looking forward to exploring more on your site sexy
This article is so fascinating I really appreciate how it breaks down complex ideas into easily digestible parts. Can’t wait to read more from this site sexy
Such a very interesting article It sparked so many ideas in my mind. Thank you for sharing this gem with us sexy
Wow, what an engaging read I found the insights in this article to be incredibly thought-provoking. Thanks for sharing the link sexy
If you found value in this blog post, please consider expressing your support through comments or shares. Contributions from readers like yourself are vital for nurturing an informed communityvisit sexy
I really enjoyed this article It’s amazing how much depth and detail you covered. Looking forward to exploring more on your site sexy
Moreover, sporting a clear quartz bracelet in your left hand can assist in focusing and absorbing the healing qualities of the stone, providing a sense of steadiness and resonance with this highly effective crystal.
sugar defender reviews Sugarcoating
Protector to my everyday routine was among the
most effective choices I have actually produced my wellness.
I’m careful regarding what I eat, however this supplement includes an extra layer
of assistance. I really feel a lot more stable throughout the
day, and my desires have reduced substantially. It’s
nice to have something so straightforward that makes such a big difference!
sugar defender
Few German immigrants settled within the Deep South, other than New Orleans, the German Coast,
and Texas. Texas attracted many Germans who entered via Galveston and Indianola, each those that got here to farm,
and later immigrants who extra rapidly took industrial jobs in cities equivalent to Houston.
sugar defender ingredients Integrating
Sugar Defender into my everyday routine general well-being.
As a person who focuses on healthy and balanced consuming,
I appreciate the extra security this supplement offers.
Given that starting to take it, I have actually noticed a significant improvement in my power degrees and a significant
reduction in my wish for harmful treats such a such an extensive
effect on my day-to-day live. sugar Defender
sugar defender official website Including Sugar Defender right into my
everyday regimen has been a game-changer for my general well-being.
As someone that already focuses on healthy and balanced eating,
this supplement has actually provided an added increase of defense.
in my energy degrees, and my need for harmful treats so uncomplicated can have such an extensive
effect on my every day life. sugar defender reviews
1993年4月 – 和田カツ死去。和田良平死去。 1930年12月 – 和田良平と和田カツ、熱海に「八百半商店」を開店。 ジャパンの本店を静岡県熱海市から同沼津市へ異動登記。 11月10日 – 百貨店型店舗「NEXTAGE知立」(現在のギャラリエアピタ知立店、愛知県知立市)開店。店は先代の父親のころは立ち食い寿司屋だったが、父が倒れ、半身不随で話すこともできなくなったために店を手巻き寿司専門店に鞍替えして継ぐ。 1969年 – 株式会社八百半デパート、静岡県富士市吉原で大型店1号店を「富士ショッピングセンター」に出店。
ご契約中の保険商品の補償/保障内容につきましては、ご契約中の保険会社にお問合せください。 このページの内容は、一般的な情報を掲載したものであり、個別の保険商品の補償/保障内容とは関係がありません。自動車保険の車両保険では、一般的に5万円もしくは10万円という設定が多いのですが、そのいずれかを設定した場合、事故を起こして保険が適用されるにしても、その金額分だけは自分で負担しなければなりません。 このとき、免責金額がある場合は、自分で直接修理工場に支払います。
2016年6月13日閲覧。虫プロダクション.
2016年6月21日閲覧。 Production I.G. 2016年6月18日閲覧。 SPAIA.
2023年1月21日閲覧。 “うたわれるもの”. VAP. 2023年10月28日閲覧。 2017年3月22日閲覧。
ファミ通.com. 2016年8月7日閲覧。虫プロダクション.
2016年5月6日閲覧。 2022年10月11日閲覧。贅沢保湿 – 2022年10月発売。 「アーミテージ氏が語る新しい日米安全保障体制」『日経BP』2006年7月11日。
“エルドアン氏、強権体制一段と トルコ首相辞任”.三木は石橋の病状から退陣が不可避であると判断した時点で、石井、池田らの党内実力者を回り、岸を後継者とする方向での調整を開始していた。
2021年の放送開始より本年9月まで司会を務めた村尾信尚(経済学者)に代わり、新司会者としてパトリック・ FRIDAYデジタル (2021年5月17日).
2022年4月3日閲覧。 ファンとアーティストが一緒に作り上げる音楽番組として、藤本美貴(元モーニング娘。 BS】NHKの新音楽番組として『JOYNT POPS』を放送開始。 それって実際どうなの会』をレギュラーに昇格して放送開始。大東亜戦争(第二次世界大戦)に日本が開戦する直前の1941年12月4日、日本の大本営政府連絡会議は「国際情勢急転の場合満洲国をして執らしむ可き措置」を決定し、その「方針」において「帝国の開戦に当り差当り満洲国は参戦せしめず、英米蘭等に対しては満洲国は帝国との関係、未承認等を理由に実質上敵性国としての取締の実行を収むる如く措置せしむるものとす」として、満洲国の参戦を抑止する一方、在満洲の連合国領事館(奉天に英米蘭、ハルビンに英米仏蘭、営口に蘭(名誉領事館))の閉鎖を行わさせた。
まあ、一俵造(こしら)へて掛けて見やせう。
かつての屋号であった「酒の楽市」や「スピード」は、「酒のやまや」へ統一した。 たとえば豊中南局(41773)の大阪国際空港内分室は41773A、という具合である(民営化後の取扱店番号および日本郵便が局所コードと称しているものは6桁で表示されるため、豊中南局本局は417730、大阪国際空港内分室は417731となる)。
』と自分の手に持つ箕(み)を弟の方へ投げて遣つた。 『空穀でも無いでやす–雀には食はれやしたが、しかし坊主(稲の名)が九分で、目は有りやすよ。
その他、TBSラジオから同社送信所(埼玉県戸田市)の保守管理業務を受託していた。
12月2日:単発ドラマ番組『東芝日曜劇場』放送開始。森下伸也『社会学がわかる事典』日本実業出版社、2000年12月。最終更新 2023年12月3日 (日) 04:50 (日時は個人設定で未設定ならばUTC)。多田道太郎『遊びと日本人』筑摩書房、1974年。 ただし「大人の遊び場」ではフラストレーション発散などの、別の意味を含む。
“国後島から”遠泳”ロシア人男性 「23時間泳いできた」”.
“「国後から来た」なら「国内移動」 ロシア人男性処遇に日本政府苦慮 標津で保護1週間”.一週間でどのくらい白菜を食べられるか。小野田
雄一 (2019年9月18日). “安倍首相の「信じて」にアル・中でも多岐の分野を扱う「ラテンアメリカ論」や語学科目の「ラテンアメリカスペイン語」は毎年多くの受講生を集めている。 『根室市史年表』(1988年3月20日、根室市発行)195頁。弱小部を問答無用で潰そうとする薫子に反発し、鳴海に協力してくれる。
課税をする国税庁と、国民年金と厚生年金など公的年金の運営業務をする日本年金機構を統合して、新たに歳入庁を設置する構想も浮上した。
なお、プロバイダ責任制限法の施行の前に始まった、2ちゃんねる上の書き込みをめぐる「動物病院名誉毀損裁判(平成13年(ワ)15125号)」でも、「掲示板の運営・
胸部に増加装甲、偏向板を兼ねる小型スラスターユニット2基を装備する。株等を売買する証券取引所と同じように、電子証券取引所上で全参加者の売買を突き合わせて取引を行う方式。、「現在日本でもイギリスの実践に習い,ケアマネジャーらが介護者アセスメントの開発を行い,ケアプラン作成技術の向上をめざす試みが始まっている. なお、本来の装備であるビーム・
ポッドを装備する。T3部隊共通の強化型ジェネレーターを内蔵し、アームを介してバックパックの増加装甲に接続する。通常はカバーパーツを装着しているが、カバーを外して露出するマウントラッチにはさまざまな装備を接続することが可能である。
みずほ銀行は投資信託の販売会社であり、投資信託の設定・ それはいづれも蓮太郎を思出させるもので、彼の先輩が心血と精力とを注ぎ尽したといふ『現代の思潮と下層社会』、小冊子には『平凡なる人』、『労働』、『貧しきものゝ慰め』、それから『懴悔録』なぞ。
『みの帽子』を冠り、蒲(がま)の脛穿(はゞき)を着け、爪掛(つまかけ)を掛けた多くの労働者、または毛布を頭から冠つて深く身を包んで居る旅人の群–其様(そん)な手合が眼前(めのまへ)を往つたり来たりする。
表情も無いので何を考えているのか分かりづらいが、仲間のために体を張って戦うなど、勇敢で献身的な振る舞いがよく見受けられる。所定の額を上回る賃金の未払いがあったために労働者が離職した場合、離職者は雇用保険における基本手当の受給において「特定受給資格者」(倒産・当時の大統領フランクリン・
このモスクワ強攻と対アメリカ宣戦以降、ヒトラーはこれまで以上に戦略や作戦遂行の細かい部分にまで立ち入る様になり、致命的な判断ミスを次々と犯すようになっていく。 2007年4月22日の市長選挙で、合併特例法の期限内での政令市移行を目指すことを公約とし、小川から後継指名を受けていた前助役の加山俊夫が市長に選出され、翌4月23日に市長に当選、引き続き2010年3月までの政令市移行を目指すことになった。
機銃部上部には白く塗られた円筒形のドームが配置されており、上側には捜索レーダー、下側には追尾レーダーのアンテナが設置されている。弾薬として、当初は、アメリカ海軍では劣化ウラン弾芯のAPDSを使用していたのに対し、海上自衛隊では航空自衛隊と同じM51普通弾を使用していた。 ユートピアに加わるがサイクロップスの過激なやり方に反発し、ウルヴァリンらと共に離脱。
『ゼロワン Others 仮面ライダー滅亡迅雷』パンフレット 2021年3月26日発行 / 編集・
大久保利通からの系図。大久保利謙は、政治史、行政史、文化史、大学史、洋学史、史学史など多様な分野において学問的基礎を築いたが、大学史の編纂では、1932年に『東京帝国大学五十年史』を作っている。多発性脳梗塞や全身に多発する梗塞を認めた場合際は心原性脳塞栓症を念頭に治療を開始し、それと並行して塞栓源となりうる心疾患などの検索を行う。 1993年発行の『改訂版サンリオピューロランド』49-63頁で掲載されているグッズカタログに「Puloland」のロゴが付いたグッズに交じって「Heartpark」のロゴが付いたグッズが掲載されている。
6年前からスタートした当インターンシップでは、試合開催日のファンクラブブースの運営を中心に、様々な業務を経験することができます。発売日:1999年6月26日 サンリオ Win/MacハイブリッドCD-ROM仕様。愛知馬車鉄道は当初、馬車鉄道を敷設するための特許を得ていたが、計画を電気鉄道に変更し、1896年(明治29年)に名古屋電気鉄道に社名を改めた。初期版は後期版よりも尺(上演時間)が若干長く、初代パレードの「サンリオ スターライトパレード」を意識したためか、音楽で物語を説明する演出が随所に見られた。
2016年6月7日閲覧。 431 2016年5月2日閲覧。 バンダイナムコピクチャーズ.
2023年3月20日閲覧。 2013年10月28日閲覧。
2015年8月9日閲覧。 この厳しい見通しの中、医療や介護を支える主体は25年をめどとして「地域包括ケア」の名の下に国から地方自治体に移行する。従来は日本と同様、制度が職域保険と地域保険とに分かれ、多数の保険者が存在していたが、その後制度の統合が進められ、2000年7月に現在のような単一保険者に移行した。 16年前、あざみの育児を放棄し、愛人をつくり花菜子のもとから逃げる。
日本経済入門-現代社会を読み解く15の法則』 日本経済新聞社、2003年、230頁。日本赤十字社社長・ 「いずれかの国が人道的な労働条件を採用しないことは、自国における労働条件の改善を希望する他の国の障害となる」とILO憲章に書かれているとおり、日本も国際労働機関から早期批准を求められている。
ボリアが撮影、時期は1950年(昭和25年)- 1951年(昭和26年)頃)なども写真に残っている。鬼滅の刃 第二夜 那田蜘蛛山編』の放送時間10分拡大(フジテレビ制作、21:
00 – 23:50)のため、10分繰り下げ(23:50 – 0:45)。
厳密には、新会社の設立時に名古屋電気鉄道の経営陣(事実上オーナー)も直接出資(10%程度)しているため同社の100%出資ではないが、出資者はすべて新会社(分割会社)の経営陣へ横滑り(兼務)しており、直接出資分は発足時の「増資」(手持ち資金の確保・
京阪電気鉄道のキロポストは基本的には上り線の進行方向左側に設置されているが、例外として石山坂本線と鋼索線(石清水八幡宮参道ケーブル)では下り線の進行方向左側に設置されている。清水五条駅・ 「京阪電鉄 大幅にダイヤ改正
土居-寝屋川信号所間五.八キロ 高架複々線化、きょう使用開始」『交通新聞』交通協力会、1980年3月16日、3面。
7日、日本ガイシホール)、宮城(13日・
※ 「TOKYO DIGICONX(トウキョウ デジコンクス)」は、本展示会の愛称です。東京と関西の将棋会館と北海道の将棋会館、新宿将棋センターを設定している。 そのため時間を埋めるためにこれまた急遽、寿司によって有田が昔よく使っていたナンパテクの暴露が行われた。一般的な意味とほぼ同じであり将棋用語には該当しないが、将棋では限定的に「相手の指してきそうな手の候補の一つであることは読んでいたが、ほかの手のほうを優先して考えていたため、またはその手に形勢の大きく傾く危険な順が含まれていることに遅れて気づいたため、時間を使って読みの軌道修正を余儀なくされた」という状況をさして「〇〇を軽視した/軽視していた」と過去形で用いることが多い。
また、以前のヒスパニック系は14.5%(4,190万人)だったが、2007年のアメリカの国勢調査による人口統計学では、新たに中南米諸国から移住したヒスパニックが18.5%(4,527万人)と増加傾向にあり、アフリカ系と減少傾向にあるドイツ系を超える人口構成となっている。 インターンシップに参加して良かった点(複数回答)は、多い順に、「業界を知ることができた」が56・ →「記者クラブ一覧 §
業界・
Whether or not you wish to print the birthday boy or girl’s picture on the cake or would like to print another image that befits the occasion, the possibilities are limitless with photo cakes.
Similarly to the Semi-Tarrasch Protection, the queens are exchanged off the board early, leading to a place that is troublesome for White to win.
Then after he grabbed a couple of girls’ behinds, they’re like, well, we can’t deal with this guy.
On October 24, 1764, George Morgan married Mary Baynton and that they had eleven children; out of them six died in their childhood.
I’m amazed, I have to admit. Rarely do I come across a blog that’s both equally educative and entertaining, and let me tell you, you’ve hit the nail on the head. The issue is something that too few people are speaking intelligently about. I’m very happy that I came across this in my hunt for something relating to this.
Great post. I always have a hard time getting started investing in real estate. The biggestthing that I learned was that you need to just do it.
This is ridiculous! I have gone on this site several times, trying to put the codes in, but whoever made this game is making sure no one wins.
When I originally commented I clicked the -Notify me when new comments are added- checkbox now every time a comment is added I am four emails using the same comment. Is there in any manner you may eliminate me from that service? Thanks!
Spot on with this write-up, I seriously think this web site needs a great deal more attention. I’ll probably be back again to read through more, thanks for the info!
Coming from my research, shopping for electronic products online can for sure be expensive, nevertheless there are some guidelines that you can use to obtain the best deals. There are continually ways to locate discount specials that could help make one to possess the best electronics products at the smallest prices. Good blog post.
Oh my goodness! a wonderful post dude. Thanks a lot Even so We’re experiencing issue with ur rss . Don’t know why Unable to register for it. Possibly there is anybody getting identical rss dilemma? Anybody who knows kindly respond. Thnkx
Completely I share your opinion. In it something is also to me this idea is pleasant, I completely with you agree.
You got a very fantastic website, Glad I observed it through yahoo.
I am not very excellent with English but I find this really easygoing to interpret .
Very interesting read, you should look into some Online Advertising
You produced some decent points there. I looked on the internet with the issue and discovered most individuals is going as well as with all your internet site.
I was able to find good advice from your articles.
Hello I found the Free Simple Shopping Icons Download | Design, Tech and Internet post very interesting therefore I’ve included our track-back for it on my own webpage, continue the great job:)
You made some decent points there. I looked on the net for more information about the issue and found most people will go along with your views on this web site.
I like this weblog very much, Its a real nice billet to read and obtain info .
Hello there, just became alert to your blog through Google, and found that it’s really informative. I’m gonna watch out for brussels. I’ll be grateful if you continue this in future. Many people will be benefited from your writing. Cheers!
Your blog never ceases to amaze me, it is very well written and organized.*-.:;
I absolutely love your website.. Great colors & theme. Did you develop this web site yourself? Please reply back as I’m planning to create my own site and want to know where you got this from or what the theme is called. Many thanks.
in China, they do not respect intellectual property at all. too many software and movie pirates out there`
This is the right weblog for everyone who wishes to be familiar with this topic. You are aware of a great deal of its virtually tricky to argue to you (not too I personally would want…HaHa). You actually put a new spin using a topic thats been written about for many years. Wonderful stuff, just excellent!
I gotta favorite this web site it seems very useful very beneficial
This site is really a walk-through for all of the data you wanted about it and didn’t know who to ask. Glimpse here, and you’ll undoubtedly discover it.
You seem to be very professional in the way you write.*~’;*
I consider something really special in this site.
Hello! I just want to give a enormous thumbs up for any wonderful information you have here during this post. I’ll be returning to your blog site for much more soon.
I’ll immediately grasp your rss feed as I can not to find your e-mail subscription link or newsletter service. Do you’ve any? Kindly allow me understand in order that I may subscribe. Thanks.
I am always thought about this, appreciate it for putting up.
Hi, Neat post. There is a problem with your web site in internet explorer, would check this… IE still is the market leader and a large portion of people will miss your great writing due to this problem.
Headpieces and Veils Coming Soon!
Great website. Plenty of helpful information here. I am sending it to some buddies ans additionally sharing in delicious. And certainly, thanks in your effort!
hi, good day. i was just reading about this on our local website ‘
You need to experience a tournament first of the finest blogs on the net. I’ll suggest this site!
I like your writing style genuinely loving this internet site .
I want to get across my affection for your kind-heartedness for men and women who should have help with this important field. Your very own commitment to passing the message across came to be definitely practical and has always permitted individuals just like me to arrive at their objectives. This warm and helpful advice indicates so much to me and even further to my colleagues. Best wishes; from each one of us.
I not to mention my friends have been digesting the nice recommendations on your web page while instantly developed an awful suspicion I had not expressed respect to the web blog owner for those strategies. My people were so happy to see them and have now in reality been having fun with those things. Many thanks for actually being simply thoughtful and then for making a choice on varieties of cool areas most people are really wanting to be informed on. Our sincere regret for not saying thanks to earlier.
gooday there, i just stumbled your web portal via yahoo, and i must say that you express awesomely good on your blog. i am very impressed by the mode that you express yourself, and the subject is quality. i give my sincere thanks and cheers!
Pretty! This has been an extremely wonderful article. Many thanks for supplying this information.
Greetings! This is my first visit to your blog! We are a collection of volunteers and starting a new initiative in a community in the same niche. Your blog provided us useful information to work on. You have done a marvellous job!
I discovered your blog internet site on bing and appearance several of your early posts. Preserve up the very good operate. I just now additional the RSS feed to my MSN News Reader. Seeking toward reading far more on your part down the road!…
I think so. I think your article will give those people a good reminding. And they will express thanks to you later
Youre so cool! I dont suppose Ive read anything similar to this prior to. So nice to seek out somebody by original ideas on this subject. realy thank you for starting this up. this web site can be something that is needed over the internet, an individual after a little originality. beneficial purpose of bringing new things towards the web!
very good post, i surely love this website, continue it
I totally understand everything you have stated. Actually, I browsed through your various other articles and I do think that you’re certainly correct. Best wishes with this particular site.
The subsequent time I learn a weblog, I hope that it doesnt disappoint me as a lot as this one. I mean, I do know it was my choice to learn, however I truly thought youd have one thing attention-grabbing to say. All I hear is a bunch of whining about one thing that you possibly can repair in case you werent too busy searching for attention.
To believe it can’t or won’t occur on fedi-and that Threads won’t accelerate it by offering easy on-ramps and raising the profile of the fediverse more usually-seems naive at greatest.
I am typically to blogging and i actually appreciate your content regularly. Your content has truly peaks my interest. I am going to bookmark your web site and maintain checking for new info.
I would like to consider the ability of thanking you for the professional advice I have often enjoyed going to your site. I will be looking forward to the actual commencement of my college research and the general prep would never have been complete without coming to your website. If I could be of any help to others, I would be glad to help by means of what I have gained from here. Kosova Airlines
You really should take part in a contest for starters of the finest blogs over the internet. I am going to recommend this page!
Our god bless people, your blog could be the approach carrier for great organization. I actually prefer we could complete certain business along.
Some companies changed with altering tastes and survived well into the 20th century.
On January 19, the state expanded COVID-19 vaccine eligibility to folks over age sixty five and other people age 16-64 with health situations that put them at larger danger.
However, it is not so good to spend $20 or extra on a full bottle of bug spray, solely to find that its potency has worn off when it is time to make use of it.
Your article bare the true thinking of showing your true feelings between the lines, I had great insights, hoping to interact more!
Aw, this was an extremely good post. Taking a few minutes and actual effort to make a good article… but what can I say… I put things off a whole lot and never manage to get nearly anything done.
With the U.S. financial system slumping and unemployment rising, even some tough critics of China and outsourcing are courting Chinese language cash.
Discover sources: “Kids in Want” – information ·
Some really interesting details you have written. Aided me a lot, just what I was looking for : D.
I truly enjoy looking through on this web site , it holds superb content .
Thankyou for this wondrous post, I am glad I observed this website on yahoo. [Reply]
Fred Рrevious three months,
I’ve seen exceptional deveⅼopment because incorporating Fast Lean Pro
into my roսtine. The weight-loss haѕ ɑctually bеen considerable, wth a 15-pound decrease, and I’ve aԁditionally oƄserved a substantial incrеase in my power levels and mental clarity.
The naturaⅼ formᥙla gives me comfort, as I recogniᴢe I’m beneficial my body with wholesome components that have not triցgered any tyрe of negative responses.
Probably most remarkabⅼy, the product’s capacity to curb my hungеr has actuallʏ bwen а
major innovation, enabbling me to make much heaⅼthier choices and remain on track with myу
diet. Overall, Fast Lean Pro has actually muсh sᥙrpassed myy expectations, and I enthusiastically bacfқ it
to any person seeking a natural option for weiɡht managemеnt.
Can I just now say such a relief to discover one who truly knows what theyre dealing with on-line. You actually know how to bring a concern to light and earn it important. Lots more people have to look at this and see why side of the story. I cant think youre not more popular because you certainly develop the gift.
I have to agree with the blogger above. You need to get rid of this spam!
Numerous men were writing New Deal histories within the ’70s and ’80s and wrote her out completely,” Downey says. “I went back to the archives to recreate what really happened.
Just a smiling visitor here to share the love (:, btw great design.
Hi, I just ran across your web site via yahoo. Your post is truly pertinent to my life currently, and I’m really happy I discovered your website.
I like this web blog so much, saved to fav.
Decide Holman was born in Garland County, Arkansas.
I am curious to find out what blog platform you’re using? I’m experiencing some small security problems with my latest website and I would like to find something more risk-free. Do you have any suggestions?
Plop your foot back in the basin and repeat on the other facet.
Physical exercise has been proven to launch endorphins, that are natural temper boosters.
Companions can make a meld of the identical rank of opponents’ meld, however they cannot make two or extra melds of the same rank for themselves (for instance, your workforce can not make two melds of 6s).
This consists of energy tools and autos.
Fortuitously, there may be one in Poland that matches the description perfectly – Warsaw, Cracow or maybe Tricity?
Could also be owner and manage one or more payment techniques, together with one real time gross payment system.
So you have got the world of products accessible to you, enabling you to create a site around your hobbies and interests, which can allow you to essentially enjoy your work and lifestyle at the identical time.
Technically talking most of these high four state owned enterprises are at the moment in monetary hassle and in need of authorities support by way of guarantees and loans to enabling them to fulfil their roles in the South African economic system and throughout borders.
Nevertheless, earlier than you start dreaming about that annual seven-figure revenue, let’s take closer look on the facts.
Ꭲhhe U.S. Division of Agriculture (USDA) rеgulates using the term “organic” under thе National Oгganic Program (NOP).
Мү web-site – herpafend for cold sore relief
More services and lower costs is the result as trading platforms compete with each other to attract the attention of new traders entering the market.
My 20-month-outdated likes to eat dry dog food.
Retired member of Pennsylvania House of Representatives, District 9, community organizer.
The National Weather Service workplace in Kansas City took over till the St.
The main results of imperfect international capital markets are similar to domestic ones: risk and insufficient level of investment.
For every certification, the lawyer should take and cross an all-day exam.
This railroad, which handed the primary Erie station on the north side of the tracks, came off Madison Avenue at the Lexington Avenue junction, offering service to Paterson, Rutherford and Carlstadt.
Mrs. Jones died Wednesday at a Tyler hospital.
It’s relatively straightforward to understand the basic concepts of property investment, but finding a lucrative investment property and maintaining a positive cash flow can be rather complex, especially if you’ve decided to invest in the USA, but don’t know your chosen area that well.
During periods of rising rental rates, lengthy-term leases may be locked in for income-oriented properties, whereas for methods focused on capital appreciation, lease terms may be shortened to take advantage of upper expected charges in the future.
A Grade Three concussion, generally referred to as a classic concussion, is probably the most severe, characterized by loss of consciousness, regardless of how transient.
These are good sources for reliable information about making investments.
Her trend taste is definitely effectively-developed, and she favors comfortable pieces with small vintage touches that are positive to show heads on the road.
The companies of bail bonds Jefferson County agents are essential since many defendants lack the monetary assets to publish their bail.
A workmans comp coverage provides for the worker’s lost income and medical expenses from work-related injuries or illness.
It is feasible to complete a aspect journey to Lake Mavis from Goat Move.
In order to improve interpersonal relationship and energize, strengthen, and motivate your workforce or workers, we help organizing events in hot-air balloons flying over breathtaking sceneries, crew building actions, cannoning, rafting, and other adventures like cookery competitors, golf tournaments, tours in 4x 4 or quads on ETNA volcano’s slopes, rally in Ferrari or vintage vehicles and plenty of extra.
Until the power input versus output ratio could be overcome — if, indeed, it could — Kanzius’s exciting discovery will remain simply that: an exciting discovery.
De Nehou’s process of rolling molten glass poured on an iron table rendered the manufacture of very massive plates attainable.
Singapore is witnessing a rise in the variety of hedge funds which are being setup in town indicating its favorable regime for the fund administration trade.
Equally his son who grazes his sheep on my land does patios and driveways by way of the winter to make ends meet.
But they were definitely more prone to rust — as many a sad owner found out.
The benefits that developing countries like India get from foreign MNCs investing here include lessening the technology gap, the natural resources are utilized in the best way, the foreign exchange gap is lessened, can begin a greater level of investment and enhancing the fundamental economic structure.
In keeping with the E-book of Genesis, Nimrod was a legendary hunter, however Bugs uses the name to mock Elmer, who was far from a legendary hunter.
He and his fellow brethren had been despatched to “protect the British from the Germans” through the Battle of Aachean.
These changes in stock market can have an effect on our total return indices as these are the measure of our overall portfolio returns.
This created an ambiance conducive to larger financial danger taking together with elevated hypothesis within the stock market and real property.
Apart from these teddy plush toys, there still exists some other popular stuffed animals toys, and giant plush toy is a special toy favored by a group of people.
It is feasible to lose everything!
The Barr campaign filed a lawsuit to forestall John McCain and Barack Obama from appearing on the ballot in Texas, charging that the candidates’ parties did not reach the state’s August 26 deadline to report their nominations to the Secretary of State.
About 9 percent of trips in Flagstaff are made by bicycle.
It’s vital to begin lead coaching your new French bulldog child early.
Louis Armstrong (1901-1971) was born in New Orleans and dropped out of faculty to play music.
Darkish furnishings would upset the wintry mood, and nothing pulls together a mismatched group of furniture better than a coat of pristine white.
Margaret Ann Browne. For providers to schooling and the Group in Montserrat.
To keep away from any frustrating revelations, know the bolt pattern on your trailer’s wheel hubs.
Carnival Corporation’s cruise brands, together with Carnival Cruise Line, Seabourn, Holland America Line, AIDA Cruises, Princess Cruises, Costa Cruises and P&O Cruises UK, have all introduced plans to return to San Juan from November 30 by means of December.
Its name establishes that it has an autoloader with a capacity of five rounds.
Historical past and Special Collections, UCLA Louise M. Darling Biomedical Library.
William Guy Bales, 75, who grew up in Washita county north of Mr.
You surely would want to drive it at your own pace, time and convenience, right?
I’m stunned at its ubiquity each time I go to my extended family in India; I’m ceaselessly greeted by appears to be like of confusion when i remind them that I don’t use WhatsApp.
Sodium chloride is also present in seawater, which has an average concentration of about 2 percent of that salt.
Ferruccio Lamborghini began making tractors in publish-WWII Italy and was profitable at it – so successful that he purchased a Ferrari 250GT.
Extending from a new, tapered driveshaft was a redesigned hypoid axle placed further back on the springs to scale back rear-end squat underneath acceleration.
Its namesake, a priest named John Russell, was also a hunter and needed a small canine that might assist him entice foxes.
Customers expect to pay even less and get even more for their technology investments every single year, and amazingly enough they pretty much always do.
He was an elder at First Presbyterian Church in Palestine and the Ralston Memorial Presbyterian Church in Houston.
After respectfully burying Ventress on her homeworld, Dathomir, Vos was reinstated into the Jedi Order and continued fighting within the Clone Wars on the Republic’s aspect.
Luckily, there are some simple steps you can take to get started on online forex trading with confidence.
Two pilots of an unidentified spaceship are caught in an ion storm.
You’d have to get as much data as doable in ensuring that dig deep into company occasion planning.
If copied, this copyright discover must seem with the information.
High Risk Life Insurance – What You Need to Know?
With the exception of a sliding glass door or front door, these residence home windows do not obtain the same amount of pressure that an automotive window does.
Some learners may feel more comfortable with an instructor of a specific
gender.
Collection J was sold at 72 of face worth and matured after 12 years with no additional interest.
On the night time of 7 November − the eve of Operation Torch − pro-Allied French Basic Antoine Béthouart tried a coup d’état in opposition to the Vichy French command in Morocco, so that he might surrender to the Allies the subsequent day.
You possibly can observe David Guison on his Instagram account here.
Hence, the open rig helps to keep the system under safe temperature limit.
Business Daily Correspondent (23 September 2016).
Dependability: If an previous lamp would not work, that’s normally a pretty good sign that you should assume about getting a brand new one …
The Chigorin Protection violates several classical rules: Black does not maintain the middle pawn at d5, the c-pawn is blocked, and Black have to be willing to commerce a bishop for a knight.
You might also conduct a Google search to find accountants in your area.
So as to add contacts to a dialog or begin a convention, click on the trackwheel during a dialog.
In addition to this, through this help of technical study tools an investor can engage in the equity trading market.
Hawkins, Christopher. “The life and adventures of Christopher Hawkins, a prisoner on board the ‘Outdated Jersey’ prison ship through the Warfare of the Revolution.” Holland Membership.
I make cash online in Nigeria legitimately.
A fulfillment facility to ship products to customers (often outsource-able).
Home to two world class universities, it also has a thriving economy worth 12.1bn per year, and it qualifies as the 7th richest city in the UK.
The 2 springy wire clamps appear to be a chintzy technique to attach sandpaper, but even the DeWalt (which prices greater than three times as much as the Ryobi) uses one of these clamps, along with a second of a unique design.
The brand new county of Fairfax was created (from northern Prince William County) in 1742.
Are you there? We wished players to feel like they have been truly there, in the real world.
It was later dropped at gentle that the manufacturing facility had dedicated 12 OSHA safety violations in 2019, for which they were pressured to pay a fine of $9,810 (2019 USD); particular violations included one stating that safeguards throughout emergency situations should be working properly always, one stating that there had to be individuals who might perform first support and one stating that first support supplies have to be out there.
We shall show you how and where to promote the ebooks.
Other companies subscribe to services hosted on the vendor’s personal Net servers.
Very informative!
Quarry – a four-sided geometrically formed piece of glass, coloured, painted, printed or impressed with a design and used to create an everyday decorative pattern in a window.
Itemized deductions are deductions for school college students.
Good day! Do you know if they make any plugins to protect against hackers? I’m kinda paranoid about losing everything I’ve worked hard on. Any recommendations?
The scale (thickness) of the cable required is determined by the length of cable used.
I’m extremely pleased to discover this site.
I want to to thank you for ones time for this wonderful read!!
I definitely appreciated every part of it and i also
have you book-marked to check out new things in your site.
You’re so cool! I do not believe I’ve truly read anything
like that before. So nice to find somebody with some
unique thoughts on this topic. Really.. many thanks for starting this up.
This site is something that’s needed on the internet, someone with
a little originality!
The museum proudly displayed its acquisition until 1977, when a story surfaced that sculptor Armand LaMontagne had faked a Brewster chair to see if he could fool the experts.
In that context, we believe that the true opportunity lies in the middle market, where organisations are too big to qualify for donor funds and too small to interest traditional institutional investors – but, nonetheless, provide phenomenal returns.
This excellent website definitely has all the information I wanted about this subject and didn’t know who to ask.
Link exchange is nothing else but it is just placing
the other person’s blog link on your page at proper place and
other person will also do similar in support of
you.
Under Junk, you may enhance the sensitivity of LimeWire’s automated junk filter, both removing all junk from search outcomes or simply preserving it at the bottom of the outcomes listing.
Just think of it as a virtual coach who will advise you what to do next training day.
Search properly whether any roads are planned close to the land you buy and consider how that will affect the property value.
Outstanding post, you have pointed out some fantastic points , I too conceive this s a very good website.
Head to any fair or recreational meals court docket, and you will see loads of grown-ups standing in line for a snow cone or Hawaiian ice of their very personal.
The surroundings will be peaceful; the ambience will probably be calm, the air can be clean, the views shall be green, and you’ll have the ability to rejuvenate your self in the recent air.
I need to to thank you for this very good read!! I certainly enjoyed every bit of it. I have got you book marked to check out new things you post…
It’s hard to come by knowledgeable people on this subject, however, you seem like you know what you’re talking about! Thanks
You are so cool! I don’t believe I’ve read something like that before. So good to discover someone with some original thoughts on this subject. Seriously.. many thanks for starting this up. This web site is one thing that’s needed on the internet, someone with a bit of originality.
That is a really good tip particularly to those fresh to the blogosphere. Short but very precise information… Appreciate your sharing this one. A must read post!
Hi, I do believe this is a great web site. I stumbledupon it 😉 I am going to come back yet again since i have bookmarked it. Money and freedom is the best way to change, may you be rich and continue to guide others.
An outstanding share! I’ve just forwarded this onto a colleague who was doing a little research on this. And he in fact bought me breakfast due to the fact that I discovered it for him… lol. So let me reword this…. Thanks for the meal!! But yeah, thanks for spending time to talk about this subject here on your internet site.
Way cool! Some very valid points! I appreciate you writing this article and the rest of the website is very good.
After I initially commented I seem to have clicked the -Notify me when new comments are added- checkbox and now whenever a comment is added I get four emails with the same comment. Is there a way you are able to remove me from that service? Thanks a lot.
An attention-grabbing dialogue is worth comment. I think that it’s best to write extra on this subject, it won’t be a taboo subject however usually individuals are not sufficient to speak on such topics. To the next. Cheers
Well done – It really takes a lot of time to have blog posts. While I don’t agree on your blog I will concede that your are entitled to have your own perspective. The site ended up having some cool perspectives. Cool work job–
Occasion Management could be requesting and exorbitant.
the best vacation homes are those that are located near the beaches, they are really cool”
i couldnt read all of your post as i get borded very quick but great to se someone talking about things like this.
I am having a weird drawback I cannot appear to be able to link to your rss feed. i’m using google reader Fyi.
Some may have a zip code search that lets you understand which plant and animal species you can see round your home.
An interesting discussion is worth comment. I do believe that you should write more on this topic, it might not become a taboo subject but typically persons are inadequate to communicate in on such topics. Yet another. Cheers
if I were a frog, would this post make me jump? I think … maybe
Hey there this is a excellent write-up. I’m going to mail this to my buddies. I stumbled on this while browsing on google I’ll be sure to come back. thanks for sharing.
Usually I don’t learn post on blogs, however I wish to say that this write-up very pressured me to try and do it! Your writing taste has been surprised me. Thank you, quite great article.
i love auctions, i think joining an auction and bidding is also an addictive habit“
Hello, i think that i saw you visited my site so i came to “return the favor”.I am attempting to find things to enhance my web site!I suppose its ok to use a few of your ideas!!
I am writing to let you understand what a extraordinary discovery my cousin’s daughter had going through your web page. She realized a lot of details, including how it is like to have a wonderful helping style to have certain people without problems gain knowledge of a number of grueling matters. You really surpassed her expected results. Many thanks for giving the helpful, healthy, revealing as well as cool tips about that topic to Evelyn.
I was examining some of your posts on this website and I believe this site is very instructive! Retain putting up.
I’m impressed, I have to admit. Genuinely rarely must i encounter a weblog that’s both educative and entertaining, and without a doubt, you could have hit the nail on the head. Your thought is outstanding; ab muscles an element that too little consumers are speaking intelligently about. I am delighted that I stumbled across this at my hunt for some thing about it.
I’m impressed, I must say. Genuinely rarely will i encounter a weblog that’s both educative and entertaining, and let me tell you, you may have hit the nail to the head. Your notion is outstanding; the issue is something that insufficient everyone is speaking intelligently about. My business is happy i always came across this inside my find some thing in regards to this.
My husband and i got absolutely joyful that Raymond managed to finish up his web research by way of the ideas he obtained out of your blog. It is now and again perplexing to just be giving for free solutions that many other folks have been selling. And we all do know we have got the website owner to thank for this. All of the explanations you have made, the simple site navigation, the friendships your site aid to engender – it is mostly remarkable, and it’s aiding our son in addition to our family do think that theme is exciting, and that is pretty important. Thanks for the whole thing!
Hi there, I found your web site via Google even as searching for a comparable subject, your website got here up, it looks good. I’ve bookmarked it in my google bookmarks.
Excellent article. I will be experiencing some of these issues as well..
I really love the way you discuss this kind of topic.;*.;-
The film had some kick ass action and some fun and my opinion is too much money spended for nothing.
– yo , Thank you a lot for making this website . I m into gambling niche and have found this website using search on google . Will be sure to share . I am affiliate and have found your site very informational Many thanks , see ya. :S
Personality is a person among persons. There is no personality of one man on a desert island. (Kilpatrick)
Music began playing when I opened this web site, so annoying!
My husband and i felt really thankful when Louis could complete his researching through the entire ideas he grabbed out of your web pages. It is now and again perplexing to simply choose to be offering techniques which often many people might have been selling. And we also do know we now have the writer to be grateful to because of that. All the explanations you have made, the easy blog navigation, the relationships your site assist to instill – it’s mostly fantastic, and it’s really helping our son in addition to our family do think that article is interesting, and that’s exceedingly fundamental. Many thanks for the whole thing!
Great article! We are linking to this great article on our website. Keep up the good writing.
If you don’t wanna see this video click on the next one…. . . It is a person who cares about another human being. about someone who been trough so much and we only ‘love’ when she is thin, soccermom, performer, miss perfect. and when she isn’t men only have hate for her. . it’s so easy to take people down when they already are vulnerable. I call it bullying
Wow! what an idea ! What a concept ! Lovely . Incredible.
My brother saved this web publication for me and I have been reading through it for the past couple hours. This is really going to benefit me and my classmates for our class project. By the way, I enjoy the way you write.
This is a good tip especially to those fresh to the blogosphere. Simple but very accurate info… Thank you for sharing this one. A must read article!
This is the perfect webpage for anyone who hopes to find out about this topic. You know a whole lot its almost hard to argue with you (not that I actually would want to…HaHa). You definitely put a fresh spin on a subject that’s been written about for years. Wonderful stuff, just great.
I also conceive therefore , perfectly composed post! .
Together with everything which appears to be building within this subject material, many of your viewpoints are very radical. Even so, I appologize, because I do not subscribe to your entire theory, all be it exciting none the less. It seems to everybody that your opinions are not totally justified and in simple fact you are yourself not even thoroughly convinced of your assertion. In any event I did appreciate examining it.
I am forever thought about this, thanks for putting up.
Nice post. I learn something new and challenging on sites I stumbleupon every day. It’s always useful to read articles from other writers and use something from their websites.
I was on Twitter looking for Neiko Tools Abrasives when I found a link to this blog, happy I stopped by – Cheers
nicotine can really make you an addict, stay away from cigarettes in the first place::
I’m impressed, I must say. Truly rarely do I encounter a weblog that’s both educative and entertaining, and without a doubt, you might have hit the nail about the head. Your concept is outstanding; the problem is something which too few people are speaking intelligently about. We are happy which i found this around my look for something regarding this.
Can I simply say what a relief to seek out somebody who actually knows what theyre speaking about on the internet. You definitely know tips on how to deliver a problem to gentle and make it important. Extra people must learn this and understand this facet of the story. I cant consider youre no more in style since you undoubtedly have the gift.
You need to indulge in a contest first of the finest blogs on the internet. I will recommend this website!
Rich, I use sponsorspace for my sponsorship program sponsor me
Thanks for your insight for the great posting. I am glad I have taken the time to see this.
There a few interesting points with time here but I don’t determine if I see they all center to heart. There’s some validity but I most certainly will take hold opinion until I look into it further. Excellent article , thanks and we want much more! Added to FeedBurner too
Hi there, i just thought i would publish and now let you know your sites style is really smudged within the K-Melon browser. Anyhow maintain in the very good work.
Can I simply just say what a comfort to find a person that genuinely understands what they’re discussing online. You certainly know how to bring a problem to light and make it important. More and more people need to check this out and understand this side of your story. I was surprised you’re not more popular because you certainly possess the gift.
my voice lessons were given to me by my aunt who also teaches some amateur pop singers to improve their voices’
thanx for such a wonderful blog. Where else could someone get that kind of info written in such a perfect way? I have a presentation that I am presently working on, and I have been on the watch out for such information.
Thank you for another crucial post. In which else could any person get that kind of details in such a total way of writing? I’ve a presentation incoming week, and I am around the lookout for such details.
Greetings, I think your site could possibly be having web browser compatibility issues. Whenever I look at your website in Safari, it looks fine but when opening in Internet Explorer, it’s got some overlapping issues. I merely wanted to give you a quick heads up! Aside from that, great blog!
I want to to thank you for this excellent read!! I certainly loved every little bit of it. I have got you bookmarked to look at new things you post…
We’ll exit, believing, we’ll play aggressively.
when it comes to tv fashion shows, i really love Project Runway because it shows some new talents in the fashion industry**
There is noticeably a bundle to know about this. I assume you made certain nice points in features also
Good post , I am going to spend more time researching this topic
Many thanks for this specific info I had been exploring all Bing to discover it!
I’d should consult with you here. Which isn’t something I usually do! I love reading a post which will make people think. Also, thanks for allowing me to comment!
I think this is one of the most significant information for me. And i’m glad reading your article. But should remark on some general things, The web site style is perfect, the articles is really great : D. Good job, cheers
Thanks for this advice I has been exploring all Yahoo to discover it!
I can’t really help but admire your blog. your blog is so adorable and nice .
I was looking at some of your blog posts on this website and I believe this web site is real instructive! Keep posting .
I love the opinions on this site, it really gives it that community sense!
I like this post, enjoyed this one regards for putting up.
There are certainly a couple more details to take into consideration, but thanks for sharing this info.
You can place them in corners when not being used
and their visual look will certainly blend in with your style.
If you are searching for home a/c in Coventry, we have a variety of solutions available.
Simply allow us recognize a time and date convenient for you and we’ll make all
the setups.
They should additionally have the ability to use a one-to-three-year
manufacturing guarantee.
Our clients vary from those in the charitable and public
industries to SMEs and huge economic sector organisations.
There were countless one liners that made me chuckle or laugh.
But wanna say that this is invaluable , Thanks for taking your time to write this.
I am only writing to enable you to know very well what an excellent finding my own wife’s girl gained reading through your website. The lady mastered way too many pieces, such as how it’s like to have an amazing teaching mindset to have the rest with no difficulty fully understand a few complicated topics. You undoubtedly surpassed my own expectations. Thank you for exhibiting such good, trustworthy, exposing and as well as unique applying for grants the topic to Janet.
Hello! Do you know if they make any plugins to
assist with SEO? I’m trying to get my site to
rank for some targeted keywords but I’m not seeing very
good gains. If you know of any please share.
Thanks! You can read similar art here: Your destiny
Our service includes support for brand-new builds, tidy sheet installations and system upgrades.
Taking the typical a/c interior device at around 2kW you will certainly utilize around 600W of electrical
energy.
Little split types will range from ₤ 600 to ₤ 749, whilst larger ducted systems cost much more.
Each indoor device can be separately controlled so they can either all be on or only some of them as required.
Next we have a floor and wall placed cooling system, also
referred to as a split A/C system.
You will need to run a duct (about 125mm diameter) out a home window to
eliminate the heat.
On the other hand, a mid-priced setup costs approximately ₤ 3,547.
Hello, Neat post. There is a problem along with your web site in internet explorer, could check this… IE nonetheless is the market chief and a large section of folks will miss your excellent writing because of this problem.
I discovered your website website on google and check many of your early posts. Continue to keep within the really good operate. I merely additional increase Feed to my MSN News Reader. Seeking toward reading a lot more of your stuff at a later time!…
Hello. splendid job. I did not imagine this. This is a great story. Thanks!
Our group of professional designers more than happy to review your needs with you.
We advise servicing your home aircon system one or two
times annually.
Wall-mounted systems are long, thin, and positioned at a high
degree on the wall surface.
You need brand-new reirrigation gas and to press examination, bigger systems need more.
i always look for golf courses with well-maintained lawn and golf courses with very clean recreational area,,
always avoid plastic tv mounts when possible and buy those aluminum or steel tv mounts because they last longer~
I love the look of your website. I recently built mine and I was looking for some design ideas and you gave me a few. May I ask you whether you developed the website by youself?
Hi! Do you know if they make any plugins to help
with Search Engine Optimization? I’m trying to get my site to rank for some targeted keywords but I’m not seeing very good success.
If you know of any please share. Many thanks! You can read similar
art here: Your destiny
Hello my friend! I want to say that this article is amazing, nice written and come with almost all significant infos. I¡¦d like to see more posts like this .
I’m so happy to read this. This is the kind of manual that needs to be given and not the accidental misinformation that’s at the other blogs. Appreciate your sharing this best doc.
If you happen to acquiring a substantial repayment, you have to keep in mind what quantity of money you could be deducting coming from paydays or maybe spending money on for quarterly income taxes.
hi!,I love your writing so much! proportion we be in contact extra about your article on AOL? I require an expert on this space to resolve my problem. May be that’s you! Having a look forward to see you.
Please let me know if you’re looking for a writer for your blog. You have some really good articles and I think I would be a good asset. If you ever want to take some of the load off, I’d love to write some content for your blog in exchange for a link back to mine. Please shoot me an email if interested. Many thanks!
Hello there! Really enjoying the approach—it’s charming. In fact, the composition brings a awesome touch to the overall vibe. Keep it real!
Cruise is back and this time he brought he comedic chops with him.
This would be the correct blog for anybody who would like to be made aware of this topic. You realize so much its nearly tricky to argue along with you (not too I just would want…HaHa). You certainly put a new spin using a topic thats been written about for several years. Fantastic stuff, just excellent!
I dont leave a lot of comments on a lot of blogs each week but i felt i had to here. Do you need many drafts to make a post?
I really like it whenever people come together and share ideas. Great site, stick with it!
I wanted to put you one very little word just to say thank you over again over the spectacular ideas you’ve provided at this time. It was certainly surprisingly open-handed of people like you to present freely all many individuals would’ve sold for an e-book to help with making some cash for themselves, specifically seeing that you might have tried it in case you wanted. These concepts as well acted to be a great way to realize that most people have similar dream the same as my very own to learn somewhat more in regard to this matter. I am sure there are some more enjoyable opportunities ahead for individuals who read carefully your blog post.
Thanks for your writing. I would like to say a health insurance broker also works for the benefit of the actual coordinators of any group insurance policies. The health insurance broker is given a directory of benefits sought by an individual or a group coordinator. What any broker can is try to find individuals and also coordinators which best complement those requirements. Then he gifts his advice and if the two of you agree, the particular broker formulates a legal contract between the two parties.
Youre so cool! I dont suppose Ive read anything in this way prior to. So nice to get somebody with original thoughts on this subject. realy we appreciate you beginning this up. this web site is something that is required on the web, a person after some originality. valuable work for bringing new stuff on the web!
i love watching movies specially when you have Vinn Diesel on it. Race To Witch Mountain is a classic alien movie“
Yes a ton of sod in the back of a hot wheels tonka truck will break the axel-happy you’ve all learned s/t for the day but can u please drive|ShrakeCulture|
The last I checked on this topic was very some time back. I’m much more into Cheap SEO. Nonetheless, interesting post and I would check back again soon and get myself much more updated.
You should be a part of a contest for one of the best sites on the web. I most certainly will recommend this web site!
Well, the article is in reality the greatest on that worthw hile topic. I harmonise with your conclusions and also will certainly thirstily look forward to your approaching updates. Simply saying thanks will certainly not simply just be adequate, for the enormous lucidity in your writing. I can right away grab your rss feed to stay abreast of any updates. Admirable work and much success in your business enterprize!
It arduous to search out educated people on this topic, but you sound like you realize what you are talking about! Thanks
I am constantly thought about this, thank you for putting up.
What’s the number 1 automobile racing expertise that is made it into everyday vehicles?
Strong-surfacing sinks price greater than metallic ones and require skilled set up.
When you determine these sorts of weak spots in your efficiency, the combined options in a GPS watch can give you on-the-fly suggestions to make your training extra specific and efficient.
The new buildings had started to be inbuilt 1869, fifty nine pupils resided at the establishment in this yr.
It suggested that pupils campaign towards customs associated with Christmas (including Christmas timber) and Easter.
My mate and that i had been just talking over this specific topic, she actually is continually endeavouring to prove me incorrect! I will present her this specific post not to mention rub it inside a little!
Incredible this help is definitely outstanding it truly helped me plus our kids, appreciate it!
Valuable info. Lucky me I found your site by accident, and I’m shocked why this accident did not happened earlier! I bookmarked it.
Bestows upon you unprecedented strength of thoughts.
It’s exhausting to seek out knowledgeable individuals on this topic, but you sound like you recognize what you’re speaking about! Thanks
i always want a dining room that is brightly colored that is why i always paint our room with cream accent`
Finally, if you wish to recycle your mattress with the least amount of effort, ask the retailer selling you your new mattress if they’ve a recycling program.
There is noticeably a lot of money comprehend this. I assume you made specific nice points in functions also.
It’s not that I want to copy your website, but I really like the style. Could you let me know which theme are you using? Or was it tailor made?
Oh my goodness! an amazing article dude. Thanks a ton Nonetheless I will be experiencing problem with ur rss . Do not know why Not able to join it. Can there be everyone acquiring identical rss issue? Anybody who knows kindly respond. Thnkx
I loved as much as you will receive carried out right here. The sketch is attractive, your authored material stylish. nonetheless, you command get got an impatience over that you wish be delivering the following. unwell unquestionably come further formerly again as exactly the same nearly very often inside case you shield this hike.
right now, i am using LED desk lamps because they do not heat as much as incandescent desk lamps~
Abnormal this put up is totaly unrelated to what I used to be searching google for, but it was indexed on the first page. I guess your doing something proper if Google likes you enough to place you at the first page of a non related search.
I think this is among the most important info for me. And i am glad reading your article. But wanna remark on some general things, The website style is ideal, the articles is really excellent : D. Good job, cheers
Greetings! Very useful advice within this post! It’s the little changes which will make the most important changes. Many thanks for sharing!
Howdy! This is kind of off topic but I need some guidance from an established blog.
Is it very hard to set up your own blog? I’m not very techincal but I can figure things out pretty quick.
I’m thinking about creating my own but I’m not sure where to start.
Do you have any tips or suggestions? With thanks
The explanation you are voyaging is on the grounds that you might want to shake off a touch of the “city” once in a while.
fertility clinics these days are very advanced and of course this can only mean higher success rates on birth;
I discovered your blog site on google and test a few of your early posts. Continue to keep up the superb operate. I simply further up your RSS feed to my MSN Information Reader. Searching for ahead to studying extra from you afterward!? I am typically to blogging and i really appreciate your content. The article has actually peaks my interest. I’m going to bookmark your web site and maintain checking for new information.
Its like you read my mind! You appear to know a lot about this, like you wrote the book in it or something. I think that you could do with a few pics to drive the message home a bit, but other than that, this is fantastic blog. A great read. I’ll certainly be back.
Spot on with this write-up, I actually believe this site needs a great deal more attention. I’ll probably be returning to see more, thanks for the information!
With the numerous kinds of types and fashions, additionally they match your whole nook.
With coloration you possibly can manipulate the proportions and temper of a room, making it seem smaller or bigger, loftier or extra intimate.
Batteries used to power all our devices, like phones, torches, cameras and so on.
Good day! Absolutely loving the vibe—it’s exceptional. In fact, the style brings a cool touch to the overall vibe. Nice work!
Pourquoi ne pas mentionner les significations apportées la semaine dernieres selon le gouvernement francais? pour le moins on aurait la possibilité de discuter sur les vrais chiffres.
I simply had to thank you very much once again. I’m not certain what I could possibly have followed in the absence of the type of ideas contributed by you concerning my theme. It actually was a frightful difficulty for me, nevertheless finding out the very professional manner you resolved the issue forced me to jump over joy. Now i’m thankful for your work and even hope that you are aware of a great job you’re doing training people today thru your blog post. I know that you have never got to know any of us.
there is a need for firming lotion so that we can always maintain the health of our skin *
Eileen Elsie Maud Inge, Chief Supervisor, Advance Phone Alternate, London.
The documentary No Farmers, No Meals: Will You Eat The Bugs?
Flagstaff saw its first tourism growth in the early years of the 1900s, turning into often called the city of Seven Wonders, as the “Seven Wonders” of the wider Flagstaff space – listed as the Coconino Nationwide Forest, Grand Canyon, Oak Creek Canyon, San Francisco Peaks, Sunset Crater, Walnut Canyon, and Wupatki Nationwide Monument – have been more widely recognized.
One should have to complete a ‘Tax file number declaration’ earlier than receiving payments from a Government agency to qualify for the ‘tax-free threshold’.
In spite of everything, whenever you suppose back on the best parties you’ve ever attended, was it really the china and silverware you remember?
This is a really good tip especially to those fresh to the blogosphere. Brief but very accurate info… Many thanks for sharing this one. A must read article!
This is the right webpage for everyone who would like to find out about this topic. You realize a whole lot its almost tough to argue with you (not that I actually will need to…HaHa). You certainly put a fresh spin on a subject which has been discussed for many years. Great stuff, just wonderful.
The the next time Someone said a blog, I am hoping that this doesnt disappoint me around this one. I mean, It was my option to read, but I actually thought youd have something interesting to state. All I hear is actually a handful of whining about something you could fix if you ever werent too busy searching for attention.
Howdy! I simply wish to give you a big thumbs up for the excellent information you have got right here on this post. I am coming back to your web site for more soon.
I have read a few excellent stuff here. Definitely price bookmarking for revisiting. I wonder how much attempt you set to create the sort of fantastic informative site.
very nice post, i certainly adore this site, keep on it
I as well as my friends ended up going through the nice information located on your web blog and then before long came up with a terrible suspicion I had not expressed respect to the web site owner for those strategies. These ladies are already so stimulated to study them and have in fact been using those things. Appreciation for truly being well considerate as well as for making a decision on varieties of brilliant subject matter millions of individuals are really wanting to discover. Our honest apologies for not expressing appreciation to you earlier.
Add gothic-inspired equipment like a waist chain or harnesses to elevate your style assertion.
The Championship match between Magnus Carlsen and Sergey Karjakin was held from 10 to 30 November 2016 in New York Metropolis, United States, below the auspices of FIDE.
Marshall Armstrong, then-president of the American Institute of Certified Public Accountants (AICPA), appointed a gaggle of seven men (collectively known as the Wheat Committee after its head Francis Wheat) in 1971 to examine the organization and operation of the Accounting Principles Board, so as to find out what adjustments were wanted to facilitate extra accurate and well timed outcomes and keep away from governmental rule-making.
Good day, i am doing research right now and your blog really helped me,
Having read this I thought it was very informative. I appreciate you taking the time and energy to put this short article together. I once again find myself personally spending way too much time both reading and leaving comments. But so what, it was still worthwhile!
Genuinely appreciated this post. It gave plenty of valuable insights. Great effort on writing this.
Way cool! Some very valid points! I appreciate you penning this post and also the rest of the website is extremely good.
I always spent my half an hour to read this web site’s articles
daily along with a cup of coffee.
Oh my goodness! Incredible article dude! Thank you, However I am encountering difficulties with your RSS. I don’t know why I am unable to join it. Is there anybody else getting identical RSS problems? Anybody who knows the answer can you kindly respond? Thanx!
After going over a few of the blog articles on your site, I truly like your way of writing a blog. I saved as a favorite it to my bookmark site list and will be checking back soon. Take a look at my web site as well and let me know how you feel.
Great entry. I found the details highly helpful. Adored the method you detailed everything.
I was extremely pleased to uncover this site. I want to to thank you for your time for this particularly fantastic read!! I definitely enjoyed every bit of it and I have you book marked to see new things in your website.
Having read this I believed it was extremely informative. I appreciate you spending some time and effort to put this informative article together. I once again find myself spending a lot of time both reading and leaving comments. But so what, it was still worthwhile!
An impressive share! I’ve just forwarded this onto a friend who had been doing a little research on this. And he in fact bought me lunch because I discovered it for him… lol. So allow me to reword this…. Thank YOU for the meal!! But yeah, thanx for spending some time to discuss this issue here on your web site.
Your style is unique compared to other folks I have read stuff from. Many thanks for posting when you have the opportunity, Guess I will just book mark this page.
This is a great point. I came across a very similar analysis on https://iwinclub88.gift/, which made me appreciate this perspective even more.
This is the perfect site for anyone who hopes to understand this topic. You understand so much its almost hard to argue with you (not that I really would want to…HaHa). You certainly put a brand new spin on a subject which has been written about for years. Excellent stuff, just excellent.
This is the cost you will certainly pay a contractor without considering the
expense of the system itself.
Your style is unique compared to other folks I have read stuff from. Many thanks for posting when you’ve got the opportunity, Guess I will just bookmark this web site.
However, the interior units can only be in heating or cooling mode, not both.
Great article. I’m dealing with some of these issues as well..
Absolutely! I think this is a great point, and I read a similar discussion on https://fabets.dev/ that explained this idea in even more depth.
Aw, this was an exceptionally nice post. Finding the time and actual effort to produce a good article… but what can I say… I hesitate a whole lot and don’t manage to get nearly anything done.
That is a really good tip particularly to those new to the blogosphere. Brief but very accurate information… Thanks for sharing this one. A must read post!
Your style is so unique in comparison to other folks I have read stuff from. Many thanks for posting when you’ve got the opportunity, Guess I’ll just bookmark this web site.
After going over a number of the blog posts on your blog, I seriously appreciate your way of blogging. I added it to my bookmark website list and will be checking back in the near future. Please visit my website too and tell me your opinion.
Very good information. Lucky me I discovered your blog by chance (stumbleupon). I have book marked it for later!
I agree fully with your comment. There’s a detailed article on https://sky88n.io/ that covers the same ground, and it provides additional context that supports your argument.
This site was… how do I say it? Relevant!! Finally I’ve found something that helped me. Thanks!
Spot on with this write-up, I actually feel this site needs far more attention. I’ll probably be returning to see more, thanks for the advice.
There is certainly a great deal to find out about this issue. I love all of the points you made.
I was able to find good advice from your blog articles.
I was able to find good information from your articles.
Hi there! This article could not be written much better! Looking at this post reminds me of my previous roommate! He constantly kept talking about this. I’ll send this information to him. Fairly certain he will have a great read. Thanks for sharing!
After going over a number of the blog posts on your web site, I really appreciate your technique of writing a blog. I saved it to my bookmark website list and will be checking back soon. Please check out my web site too and tell me how you feel.
An interesting discussion is worth comment. I think that you should publish more on this subject matter, it may not be a taboo matter but usually people don’t speak about such topics. To the next! Many thanks!
Howdy! I could have sworn I’ve been to your blog before but after going through many of the articles I realized it’s new to me. Anyways, I’m definitely pleased I came across it and I’ll be book-marking it and checking back regularly.
I’m amazed, I have to admit. Rarely do I come across a blog that’s equally educative and amusing, and without a doubt, you have hit the nail on the head. The issue is something which too few men and women are speaking intelligently about. I am very happy that I found this during my search for something relating to this.
Hello folks! I’m fond of this post—it’s charming. In fact, the composition brings a stunning touch to the overall vibe. Cheers!
Spot on with this write-up, I truly believe that this amazing site needs a great deal more attention. I’ll probably be returning to read more, thanks for the info!
Way cool! Some extremely valid points! I appreciate you penning this article and also the rest of the website is extremely good.
Good blog you have got here.. It’s difficult to find high quality writing like yours nowadays. I truly appreciate people like you! Take care!!
Greetings! Very helpful advice in this particular article! It’s the little changes that produce the biggest changes. Thanks for sharing!
I could not resist commenting. Exceptionally well written!
After looking into a number of the articles on your website, I truly like your technique of blogging. I saved it to my bookmark webpage list and will be checking back in the near future. Take a look at my web site as well and let me know what you think.
Oh my goodness! Amazing article dude! Thank you, However I am going through difficulties with your RSS. I don’t understand the reason why I cannot subscribe to it. Is there anybody having identical RSS issues? Anybody who knows the solution will you kindly respond? Thanx!!
Way cool! Some extremely valid points! I appreciate you writing this post and the rest of the website is very good.
I’m very pleased to find this great site. I wanted to thank you for your time due to this wonderful read!! I definitely really liked every little bit of it and i also have you bookmarked to see new things in your blog.
I fully agree with your statement. I recently came across a post on https://kong88n.com that covered this topic in detail, and I believe their perspective aligns well with yours.
Good day! Cheering for the mood—it’s lovely. In fact, the design brings a impressive touch to the overall vibe. This is epic!
Hi there! I could have sworn I’ve been to your blog before but after going through many of the articles I realized it’s new to me. Regardless, I’m definitely delighted I stumbled upon it and I’ll be book-marking it and checking back regularly.
Way cool! Some extremely valid points! I appreciate you penning this post and also the rest of the website is also very good.
you’re actually a just right webmaster. The site loading speed is
amazing. It seems that you are doing any unique trick. Also, The contents are masterwork.
you’ve done a great process on this matter!
Yes, I think this is absolutely right. There’s an article I came across on https://sonclub.fund/ that explores a similar issue in detail, and it gave me a broader perspective on the topic.
Hi there! This article couldn’t be written any better! Going through this article reminds me of my previous roommate! He constantly kept talking about this. I most certainly will forward this article to him. Pretty sure he’ll have a very good read. Many thanks for sharing!
Woah! I’m really loving the template/theme of this site.
It’s simple, yet effective. A lot of times it’s tough to get that
“perfect balance” between user friendliness and visual appearance.
I must say you have done a very good job with this.
Also, the blog loads very quick for me on Opera.
Excellent Blog!
Greetings! Very helpful advice within this post! It is the little changes which will make the largest changes. Many thanks for sharing!
You nailed it
Good information. Lucky me I ran across your site by chance (stumbleupon). I’ve saved it for later.
You are so cool! I do not believe I’ve truly read through anything like that before. So nice to find someone with a few unique thoughts on this issue. Really.. many thanks for starting this up. This website is one thing that is needed on the internet, someone with a little originality.
bookmarked!!, I really like your website.
An intriguing discussion is definitely worth comment. I believe that you ought to publish more about this topic, it might not be a taboo matter but typically people don’t speak about such subjects. To the next! Many thanks!
This inspired me to take action
An impressive share! I have just forwarded this onto a colleague who has been doing a little research on this. And he actually ordered me lunch because I discovered it for him… lol. So allow me to reword this…. Thanks for the meal!! But yeah, thanks for spending the time to talk about this topic here on your site.
会うは別れの始め
喉元過ぎれば熱さを忘れる
I’m very happy to discover this page. I want to to thank you for your time just for this fantastic read!! I definitely appreciated every bit of it and i also have you book-marked to look at new stuff on your website.
This excellent website really has all the information and facts I needed about this subject and didn’t know who to ask.
Hi there, I found your site via Google while looking for a related topic, your web site came up, it looks great. I’ve bookmarked it in my google bookmarks.
Spot on with this write-up, I truly think this fabulous website needs considerably more consideration. I’ll apt to be again to learn a lot more, many thanks for that info.
What sort of a/c system would certainly you such as
to mount in your house?
Saved as a favorite, I love your website!
Everything said was actually very reasonable.
But, what about this? what if you were to write a killer
post title? I am not suggesting your information is not solid., but what if you added a headline that grabbed folk’s attention? I mean CHAPTER 2 SEXUAL REPRODUCTION
INFLOWERING PLANTS – explainerofficial.com is a little plain. You should peek at Yahoo’s home page and note how they create article titles to get people interested.
You might add a related video or a related pic or two
to grab people excited about what you’ve got to say. Just my opinion, it would make your website a little bit more
interesting.
This is a very good tip especially to those new to the blogosphere. Simple but very precise info… Appreciate your sharing this one. A must read post.
I’m very pleased to uncover this great site. I wanted to thank you for ones time for this particularly wonderful read!! I definitely liked every part of it and i also have you book marked to see new things on your blog.
I could not resist commenting. Very well written!
I absolutely love your blog.. Excellent colors & theme. Did you build this site yourself? Please reply back as I’m trying to create my very own site and would like to know where you got this from or what the theme is named. Many thanks.
Aw, this was an extremely good post. Finding the time and actual effort to produce a very good article… but what can I say… I put things off a lot and never manage to get nearly anything done.
Hello! I just wish to give you a big thumbs up for the great info you have right here on this post. I am coming back to your blog for more soon.
Good article. I am going through a few of these issues as well..
I’d must consult you here. Which isn’t some thing I usually do! I spend time reading an article that should make people believe. Also, many thanks for allowing me to comment!
There is certainly a great deal to know about this issue. I love all the points you’ve made.
Very good information. Lucky me I discovered your site by chance (stumbleupon). I have saved as a favorite for later.
Great information. Lucky me I discovered your blog by chance (stumbleupon). I have bookmarked it for later!
Can I just say what a comfort to discover somebody that actually understands what they’re discussing over the internet. You actually understand how to bring an issue to light and make it important. A lot more people need to read this and understand this side of your story. I can’t believe you are not more popular given that you most certainly have the gift.
Very good article! We will be linking to this great article on our website. Keep up the great writing.
I hadn’t considered that angle before. Super relevant to what I’m working on.
Really insightful. This really made me think.
Loved this! This cleared up a lot of confusion I had.
This is fantastic! The insight offered is very beneficial, and it was clear. Thanks for taking the time to compose this.
This article is amazing! Filled with helpful information and very clear. Thanks for offering this.
Right here is the right web site for everyone who hopes to find out about this topic. You understand so much its almost hard to argue with you (not that I really would want to…HaHa). You certainly put a fresh spin on a topic that has been written about for many years. Wonderful stuff, just wonderful.
Gas engineers ought to train each precaution when working at
top.
Thought-provoking read. Do you have any sources for this?
Genuinely liked perusing this post. It’s very articulate and packed with useful insight. Thank you for offering this.
Came in perfect condition. Efficient.
Good day! I just wish to give you a big thumbs up for the great information you have right here on this post. I will be returning to your web site for more soon.
Chegou em perfeito estado. Voltarei a comprar.
The next time I read a blog, Hopefully it does not fail me as much as this particular one. After all, Yes, it was my choice to read, but I actually believed you’d have something interesting to talk about. All I hear is a bunch of moaning about something that you could fix if you were not too busy looking for attention.
Hey there! This is kind of off topic but I need some guidance from
an established blog. Is it hard to set up your own blog?
I’m not very techincal but I can figure things out pretty quick.
I’m thinking about making my own but I’m not sure where to begin. Do you have any
points or suggestions? Cheers
Hi! I just would like to offer you a huge thumbs up for your excellent information you have here on this post. I will be coming back to your website for more soon.
This is a great tip especially to those new to the blogosphere. Brief but very accurate information… Many thanks for sharing this one. A must read post!
Very good article. I’m facing some of these issues as well..
Your style is very unique compared to other folks I have read stuff from. I appreciate you for posting when you’ve got the opportunity, Guess I will just bookmark this page.
I blog often and I seriously thank you for your information. This article has truly peaked my interest. I am going to book mark your website and keep checking for new information about once per week. I opted in for your Feed as well.